Die Evolutionstheorie, ihre Bedeutung und ihre Grenzen
Zusammenfassung
Einleitend werden die Methoden der Naturwissenschaften und ihre Grenzen dargestellt. Nach einem kurzen historischen Überblick über den Weg zu Darwin wird sein Werk mit seinen Verdiensten in der Entdeckung der Variation der Arten und ihre Selektion durch die natürliche Zuchtwahl („Kampf ums Dasein“, „Überleben der Tüchtigsten“1) und die moderne Evolutionstheorie aufgezeigt einschließlich der Problematik der Abstammung des Menschen und seiner Sonderstellung in der Natur. Dann werden Alternativen zur Evolutionstheorie wie das Intelligent Design Konzept und die Schöpfungswissenschaft umrissen. Die gesellschaftliche Bedeutung des Darwinismus als Basis des Marxismus und der Rassenideologie sowie die Frage der Vereinbarkeit mit dem Christentum werden diskutiert. Die Arbeit schließt mit einer Empfehlung für einen realistischen Standpunkt eines Naturwissenschaftlers zur Evolutionstheorie.
Schlüsselwörter: Darwinismus, „Kampf ums Dasein“, Variabilität, Selektion, Abstammung, Intelligent Design, Schöpfungswissenschaft, Marxismus, Rassenideologie, Christentum
Abstract
First the methods of sciences and their limitations are presented. After a short historical review of the way to Darwin, his work and his merits in discovering the variation of species and natural selection, (“struggle for life”, “survival of the fittest”1), and modern evolution theory are shown including the problem of the descent of man and his exceptional place in nature. Then alternatives to evolution theory are outlined like Intelligent Design and creation science. Concerning the human society, the importance of Darwinism as the basis of Marxism and racism is discussed as well as the question of its compatibility with Christianity. The paper is concluded with the recommendation of a scientists for a realistic attitude towards evolution.
Keywords: Darwinism, “Struggle for Life”, Variability, Selection, Descent, Intelligent Design, Creation Science, Marxism, Russism, Christianity
Einleitung: Über die Methoden in den Naturwissenschaften
„… Evolution by natural selection is a theory, not a fact…“ – By the University board of Regents (2006)
Dieser zitierte Satz stand auf dem Aufkleber einer Ausgabe des internationalen Wochenjournals der Naturwissenschaften „Nature“.
Eine Theorie steht als die durch gedankliche Betrachtungs- und Erfahrungsweise gewonnene Erkenntnis im Gegensatz zu dem durch Erfahrung gewonnenen Wissen, wie z. B. im reproduzierbaren Experiment. Eine Theorie gilt als Meilenstein im Fortschreiten der Forschung, ist aber stets, historischer Abwandlung entsprechend, neu sich auftuenden Erkenntnishorizonten unterworfen.
150 Jahre intensiver Forschung im Geiste Darwins reichten offenbar nicht aus, um die Evolutionstheorie Tatsache werden zu lassen, wie aus dem Zitat gefolgert werden kann. Die Publikationen über die Deutung der zahlreichen Ergebnisse vor allem der Paläontologie, Archäologie, Geologie und der Molekularbiologie reichen von einer strengen materialistisch-atheistischen in der Evolutionstheorie bis zu den Alternativen, die gemäß der Informationstheorie einen intelligenten Schöpfer fordern.2
Welche Methoden stehen dem Naturforscher grundsätzlich zur Verfügung? Dieser wird immer, wenn möglich, das Experiment, die Methode der exakten Induktion, wählen. Sie wurde von Galileo Galilei erstmals bei der Aufstellung der Fall- und Pendelgesetze angewendet, was ihm den berechtigten Ruf des Begründers der naturwissenschaftlichen Methode einbrachte. Diese prüft auf Grund von Beobachtungen ein hypothetisch angenommenes Gesetz, in dem ein deduktiv abgeleiteter konstruierter Fall im Versuch erprobt wird. Dabei werden Gesetzeszusammenhänge aufgedeckt und Voraussagen gestattet, so dass im Prinzip die induktive Prüfung eines einzigen deduktiv abgeleiteten Einzelfalles genügt, um einen zwingenden Beweis zu erhalten. Der Physikochemiker Max Thürkauf nennt dies die „systematisch reproduzierbare Hervorbringung des Gegenstandes“. Der Geologe und Biologe Joa-chim Scheven stellt heraus: „Eindeutig wahr ist nur der durch das Experiment erhärtete Kausalschluss oder der Direktbeweis. Er ist Grundbaustein jedes logisch-deduktiven Gedankengebäudes und das eigentliche Werkzeug der Vernunft.“3 Diese Methode hat vor allem der Physik und der Chemie und den experimentellen Disziplinen der Biologie große Erfolge gebracht. Sie bildet die große Stärke der Naturwissenschaften und wurde Grundlage der Computertechnik, der Weltraumfahrttechnik und der Kunststoffchemie.
Die historischen Naturwissenschaften, wie Geologie und Paläontologie, arbeiten nach der zweiten naturwissenschaftlichen Methode, der generalisierenden Induktion. Diese ist eine vergleichende Methode, die Ähnlichkeiten und Verschiedenheiten der genau beschriebenen Naturobjekte und ihrer Teile und die sich vollziehenden Prozesse vergleicht und in ein System von allgemeinen Begriffen und Aussagen bringt.4 Über klassifizierende Kennzeichnung und Ordnung können fundamentale Beziehungen und kausale Abhängigkeiten aufgedeckt werden. Aber da der generalisierenden Induktion das Experiment nicht zur Verfügung steht, ist nur eine indirekte Erschließung von Zusammenhängen möglich. Der Vergleich noch so vieler Fälle kann einen Beweis nicht sicher erbringen. Man kann allenfalls von Regeln großer Wahrscheinlichkeit sprechen. Diese Methode gilt also für alle historischen Vorgänge, die sich direkter Beobachtung entziehen oder in einem Experiment nicht reproduzierbar sind und daher nur den Charakter hypothetischer Aussagen haben, so einsichtig Schlussfolgerungen aus sorgfältig zusammengestelltem Fundmaterial auch sein mögen. Da die Evolutionslehre eine historische, naturwissenschaftliche Theorie ist, bedeutet dies, dass z. B. alle „Stammbäume“ mit der Anordnung der Organismen in einem System nach abgestufter Ähnlichkeit zwar immer in einem Entwicklungszusammenhang gesehen werden können, aber immer hypothetisch bleiben.
Nach Francis Crick, der zusammen mit Watson den Nobelpreis für die Konstruktion des DNA-Doppelhelixmodells erhielt, sind die Naturforscher aufgefordert, die Naturgesetze nach Maßstäben von Atomen und Molekülen zu erforschen, d. h. eine materialistische Erklärung anzustreben. Bei diesem methodischen Atheismus setzen Naturforscher unbewusst und notwendig einen Schöpfer voraus, der die Einrichtungen und die erforschbaren Gesetze einer natürlichen Offenbarung geplant und geschaffen hat (Max Planck), die es nachzuarbeiten gilt, die auch die eigene Denkfähigkeit ermöglichen. Diese wird nicht selten als ein Lobpreis auf den Schöpfer empfunden. Ein bewegtes Staunen hat immer eine Rolle gespielt. Nobelpreisträger und Physiker Max von Laue bemerkt: „Dass wissenschaftliches Wahrheitserlebnis in irgendeinem Sinne Gottesschau ist, dürfte gerade den Besten unter ihnen (Naturwissenschaftlern) aus dem Herzen gesprochen sein.“
Die Evolutionslehre als Hypothese hat dennoch einen hohen Wert; denn allgemein hat eine Hypothese auch dann einen Wert, wenn sich später herausstellt, dass sie falsch ist. So konnte J. Dalton in seiner Atom-Hypothese davon ausgehen, dass die chemischen Elemente aus kleinsten, nicht weiter zerlegbaren Teilchen, den Atomen (atomos, gr. unteilbar), bestehen. Auf Daltons und Avogadros auf Moleküle erweiterte Molekular-Hypothese wurden z. B. die kinetische Gastheorie und die Gasgesetze, wie das Boyle-Mariottesche Gesetz, aufgebaut.5 Später aber stellte sich heraus, dass diese Hypothese nur eine grobe Annäherung an die Wirklichkeit, d. h. eigentlich falsch war. So sind Hypothesen notwendige, wichtige Schritte auf dem Weg zu vertiefter Naturerkenntnis. Sie sind zur Erprobung an eine neue Methode geknüpft. Gelingt es, eine neue Methode zu finden, gilt der Satz des Nobelpreisträgers Adolf Butenandt: „Eine neue Methode bedeutet einen Fortschritt in der Naturerkenntnis“.6 So ist auch die Evolutionstheorie als Meilenstein zu einer vertieften Naturerkenntnis zu betrachten. Der Baseler Zoologe Adolf Portmann hatte gefordert, die Evolutionslehre auf die Stufe der Hypothese zurückzuversetzen, von wo aus vielseitige Anregungen und fruchtbare Impulse für die Forschung möglich sind.7
1. Zur Vorgeschichte der Lehre von Darwin
Die Gedanken zur Herkunft der darwinschen Entwicklungslehre hat Ilse Jahn in einem umfangreichen Werk zusammengetragen.8 Im Folgenden sollen die wichtigsten Erwähnung finden.
Als der Mensch zum fest siedelnden Ackerbauer und Tierzüchter wurde, war er in die Lage versetzt worden, durch gezielte Züchtung der Tiere und Kultivierung der Nutzpflanzen die notwendigen und optimalen Bedingungen zu beachten und anzuwenden. Die ältesten nachgewiesenen Landwirtschaftszentren im Jungpaläolithikum vor 13 000 Jahren liegen im Fruchtbaren Halbmond des Nahen Ostens.9 Die Landwirte und Gärtner und später die Forstleute schufen die Grundlagen für die heutige agrarbiologische Wissenschaft und machten von der Selektion geeigneter Varianten von Tier und Pflanzen Gebrauch. Dafür seien ein paar Beispiele angeführt.
Der Haushund in seinen vielen Rassen stammt von Wolfs- und Schakalarten ab und wurde vom Menschen für seine Zwecke z. B. als Hütehund oder Jagdgefährte gezüchtet. Verwildern die Hunde, so verlieren sie die vom Menschen angezüchteten Eigenschaften, wie Hängeohren und gerollte Schwänze.
Die Kartoffel, die nach der Entdeckung Amerikas in zahlreichen Kultursorten als Werk indianischer Ackerbauern vorgefunden wurde, war bereits Hauptnahrungsmittel der Inkas (Peru), der Mayas (Guatemala) und der Azteken (Mexiko). Sie trat in Hungersnöten auch in Europa ihren Siegeszug an.
Siegismund Markgraf aus Berlin stellte 1747 fest, dass sich Zucker aus der Runkelrübe Beta vulgaris gewinnen ließ. 1802 baute Achard die erste Zuckerfabrik in Kunern/Niederschlesien. Der Zuckergehalt konnte von 2 bis 3% auf 17%, dann sogar auf 27% gesteigert werden. Die Zuckerrübe trat in der Zeit der Kontinentalsperre unter Napoleon, als die Lieferung von Zuckerrohrzucker nicht möglich war, ihren Siegeszug an.
Unter den Vordenkern des Entwicklungsgedankens findet man den großen Denker des Altertums Aurelius Augustinus (354 bis 430) aus Thagaste. Er erkannte bereits sowohl die Veränderlichkeit als auch die Konstanz der Lebewesen und konstatierte, Gott habe die Objekte in der Natur in unterschiedlichen Zuständen erschaffen, einige vollständig, die anderen in einem „potenzialen latenten Zustand, die auf günstige Bedingungen der Umwelt zur vollen Entfaltung warteten“. Sie verfügen über Potenzen, die fähig sind zur weiteren Entwicklung gemäß eines Plans; denn nie könne eine solche Entwicklung beliebig verlaufen. Bei den meisten Biologen der Zeit vor Darwin herrschte die Vorstellung einer im wesentlichen Artkonstanz, die ja Voraussetzung des Bestimmens und Wiederfindens von Organismen sowie der Vererbung ist. Doch da die Entwicklungsvorgänge jedem Biologen bekannt waren, wie die Keimesentwicklung und das Wachstum, kam auch frühzeitig der Gedanke an Werdeprozesse bei der Entstehung der Organismenvielfalt auf. Niels Stensen (1638 – 1686), auch Nikolaus Steno genannt, entdeckte das für die Abstammungslehre wichtige Lagerstättengesetz: Bei der Ablagerung der Sedimente folgt Schicht auf Schicht, so dass die jüngere Schicht auf der älteren liegt. Bei ungestörter Lagerung geht die Richtung vom Liegenden zum Hängenden, dem die Reihenfolge vom Älteren zum Jüngeren entspricht. Hier war ein wichtiger Schritt von den gegenwärtig lebenden Organismen zu den in den Sedimenten verborgenen früher gelebten Organismen getan und man konnte auch die Folge ihres Daseins ablesen. Auch konnten die gegenwärtigen Organismen mit den früheren verglichen und beide in einem gemeinsamen System untergebracht werden. Carl von Linné (1707 – 1778) stellte sich die Frage, wie man die ungeheure Organismenvielfalt in einem naturgemäßen System ordnen und wie man einen entdeckten Organismus darin einordnen kann. Linné unterschied drei Naturreiche: Pflanzen-, Tier- und Mineralsystem. Sein besonderes Verdienst ist ein neues Pflanzensystem mit 21 bzw. 24 Klassen seines späteren Sexualsystems einer neuen Bestimmungsmethode nach Anzahl und Struktur der Blütenorgane. Sein Hauptwerk ist „Systema naturae“ 1735. Linnés Verdienste liegen neben der Klassifizierungsmethode in der Festlegung einer Merkmalsterminologie und der binären Nomenklatur mit Art- und Gattungsnamen, die im Prinzip bis heute gelten. Die erste systematische Ordnung in seiner Neuauflage enthielt auch die Herrentiere – Primaten –, zu denen Linné den Menschen in vier Varianten stellte. Er hielt wohl als Systematiker an der Artkonstanz fest.10
Immanuel Kant (1724 – 1804) hatte sich naturphilosophisch sehr treffend geäußert. Er schrieb von den verschiedenen Rassen des Menschen: „Die Vorsorge der Natur… ist bewundernswürdig und bringt bei der Wanderung und Verpflanzung der Tiere und Gewächse, dem Scheine nach, neue Arten hervor, welche nichts anderes als Abartungen und Rassen von derselben Gattung sind, deren Keime sich nur gelegentlich in langen Zeitläufen auf verschiedene Weise entwickelt haben“. Heute gebührt Kant für seinen Scharfsinn Bewunderung, denn die Variationen der Arten, Rassen oder Gattungen sind bis heute die einzig beobachtbaren Taxa geblieben. Jean-Baptiste de Lamarck (1744 – 1829) lehrt zwar noch die Urzeugung vor allem von primitiven Organismen der „generatio spontanea“, aus der sich Tiere und Pflanzen entwickelt haben sollen. Die Urzeugung wurde von Louis Pasteur (1822 – 1895) widerlegt: „Omne vivum ex vivo“. Moderne Hypothesen, wie die von Stanley Miller (* 1930), gehen von der Annahme aus, dass vor der Urzeugung in einer Ursuppe mit Uratmosphäre in einer chemischen Evolution organische Substanzen entstanden sind, die schließlich zu replikationsfähigen Molekülen, wie den Nukleinsäuren, führen könnten. Lamarck lehrte die Umwandelbarkeit der Arten in großen Zeiträumen. Bei jedem Tier bewirkt der konstante „Nichtgebrauch eines Organs, dass dasselbe schwächer wird und schließlich verschwindet“, während der „häufige Gebrauch des Organs dasselbe schließlich stärkt“. Alles, was die Individuen durch den Einfluss der Verhältnisse erwerben oder verlieren, wird auf die Nachkommen durch die Fortpflanzung vererbt, wie die Giraffe, deren Hals immer länger wird, damit das Laub der hohen Baumäste erreichbar werden kann oder der Grottenolm, dessen Augen wegen des ständigen Lebens in Dunkelheit nicht mehr gebraucht werden und sich daher zurückbilden. Diese Annahme der Vererbung erworbener Eigenschaften hat sich bis nach Darwin erhalten und wurde von dem sowjetischen Agrarbiologen Trofim Denisovitsch Lyssenko (1898 – 1976) aufgegriffen. Lamarcks Evolutionstheorie ist wesentlich auf einer Stufenleiter gegründet. George Cuvier (1768 – 1832) schuf die Katastrophentheorie, die er als Begründer der Paläontologie, die sich mit der Erforschung der Fossilien befasst, vertrat. Nach seiner Theorie wurde die Erde immer wieder von Katastrophen heimgesucht, nach denen durch Schöpfung die Lebewesen neu entstanden. Eine solche Katastrophe war auch die Sintflut. Die Lebewelt der Vergangenheit zeigt eine korrelierbare Ordnung in ihrer Abfolge. Heute unterscheidet man neben vielen kleineren 5 große globale Katastrophen der Erdgeschichte („BIG FIVE“) mit einem Artensterben von 75 – 95% in Mio. Jahren jeweils vor heute: Ende Kreide 65, Ende Trias 200, Ende Perm 251, Spätedevon 376, Ende Ordovicium 443.11 Das massenhafte Aussterben wird auf Zusammenbrüche der Ökosysteme zurückgeführt, die möglicherweise eine Folge des Aufpralls von Asteroiden oder sehr großen Meteoriten auf die Erde sind. Nach jeder Katastrophe entstehen neue Typen, die vorher nicht gefunden werden konnten. Die heute bekannte kambrische Explosion vor 600 Mio. Jahren zeigt die Entstehung fast aller Baupläne der Tiere. Seitdem sind keine neuen Tierstämme entstanden. Die Triassische Auslöschung der Organismenwelt vor 200 Mio. Jahren hat eine folgende Entfaltung der Reptilien gebracht, bis diese Ende der Kreidezeit vor 65 Mio. Jahren fast wieder bis auf Reste verschwanden, worauf die Entfaltung der Säugetiere folgte. Heute hat die Katastrophentheorie wieder an Bedeutung gewonnen. So hat der Ausbruch des bis 1980 ruhigen Vulkans Mt. St. Helens sehr rasche Sedimentationen und die Bildung eines 30m tiefen Canyon in kurzer Zeit gezeigt.12 Von großer Bedeutung für die Selektionstheorie Darwins war die Streitschrift des Nationalökonomen und Sozialphilosophen Thomas Robert Malthus (1766 – 1834) „Essay on the Principle of Population“. Seine bekannten Sätze sind: 1. Die Bevölkerung ist notwendig durch die Unterhaltsmittel begrenzt. 2. Sie wächst da stetig, wo sich die Unterhaltsmittel vermehren, es sei denn, sie werde durch sehr mächtige und offensichtliche Hemmnisse gehindert. 3. Die Hemmnisse und jene, welche die übermächtige Zeugungskraft zurückdrängen, lassen sich alle in sittlicher Enthaltsamkeit, Laster und Elend zusammenfassen. Malthus behauptet, die Bevölkerung verdopple sich bei unbeschränkter Fortpflanzungsmöglichkeit alle 25 Jahre. Die Bevölkerung vermehre sich nach einer geometrischen Progression 1, 2, 4, 8,…, dagegen wachse die Nahrungsmittel nur in einer arithmetischen Progression 1, 2, 3, 4… (Malthusianismus). Diese starke Vermehrung einer Population wurde für Darwin die entscheidende Basis für die Selektionstheorie durch natürliche Zuchtwahl (Der Neue Herder 1949). Charles Lyell (1797 – 1875) schrieb die „Principles of Geology“, in dem das aktualistische (uniformitaristische) Prinzip vertreten wurde. Nach diesem sollen die geologischen Prozesse der Vergangenheit mehr oder weniger denen der Gegenwart gleichen. Dieser Aktualismus beeinflusste Darwin in seinen paläontologischen Nachweisüberlegungen für die Entstehung der Arten. Nach Lyell sterben die Organismen aus, sobald sich die Umwelt verändert und neue Organismen treten an ihre Stelle. Der Begründer der Tiergeographie ist Alfred Russel Wallace (1823 – 1923). Er untersuchte auf vielen Reisen die Verbreitung der Tiere und teilte die Erde in tiergeographische Zonen ein, vor allem in eine alt- und neuweltliche Fauna. Die Trennung zwischen den Regionen wurde Wallace-Linie genannt: Sie verläuft zwischen Borneo, Celebes und Bali in Nord-Süd-Richtung. Unabhängig von Darwin vertrat Wallace die Veränderlichkeit der Arten. Seine Arbeit „On the Tendency of Varieties to Depart Indefinitely from the Original Type" war im Jahre 1858 erschienen und ermutigte und regte Darwin an. Als Darwins Konkurrent konnte er aber für die Entstehung der Arten keine Erklärung bieten. Nur wenig später als Darwin publizierte der Augustinerpater Gregor Mendel (1822 – 1884) im Jahre 1865 seine Versuche über die Pflanzenhybriden. Jedes Merkmal wird unabhängig und ohne sich zu „vermischen“, also konstant, von den Stammformen auf die Nachkommen vererbt. Seine nach ihm benannten genetischen Gesetze fanden so wenig Interesse, dass sie erst um 1900 von Carl Erich Correns (1864 – 1933), Erich von Tschermak-Seysenegg (1871 – 1962) und Hugo de Vries (1848 – 1935) wieder entdeckt werden mussten. Mit Mendel begann die wissenschaftliche Vererbungslehre, die Darwin fehlte. Seine Ergebnisse zeigten die Konstanz der Vererbung und bestätigten im Prinzip eine Konstanz der Arten. Bis Darwin galt allgemein, dass jede Art unabhängig voneinander erschaffen wurde mit Eigenschaften, die in eine vorher bestimmte Umwelt passte. Auch Darwin trat seine Forschungsreise mit dieser Überzeugung an.

2. Darwins Werk (Abb. 1)
Charles Robert Darwin (1809 – 1882) schuf die Grundlagen der modernen Evolutionstheorie. Die Forschungsreise auf der Beagle führte ihn u. a. nach Südamerika, zu den Galapagos-Inseln, nach Tahiti und Neuseeland. Sie bedeutete einen Wendepunkt in Darwins Leben nach einem abgeschlossenen Theologiestudium und anderen Studien. Dabei hatte ihn die aktualistische (uniformitaristische) Geologie Lyells in der Forschung stark angeregt. Darwin bewunderte die Natur, unter der er die Wirkung und Leistung vieler Naturgesetze verstand, welche die nachgewiesene Aufeinanderfolge der Ereignisse bewirken. Die südamerikanischen Fossilien ließen erkennen, dass die heute lebenden Arten mit den ausgestorbenen Arten verwandt waren, also als Stammformen von heutigen Formen gesehen werden können. Darwin erkannte auf den Inseln die geographische Isolation als Basis für die Ausprägung bestimmter Inselarten und das Verdriften von Samen und Insekten auf einsamen Inseln. Ganz wesentlich waren für Darwins Weg die vielen Kontakte zu Tier- und Pflanzenzüchtern, die ihn von der großen Variabilität der Arten überzeugten. Durch Korrespondenz mit Fachleuten versuchte er über Tauben, Enten, Kaninchen und Pflanzen Bestätigung seiner Gedanken zu finden. Studien von Florenkatalogen führten ihm die weite Verbreitung der Varietäten und Gattungen vor Augen. Bereits 1837 hatte er sich eine Theorie des Artenwandels erarbeitet. Doch wie war dieser möglich? Darwin postulierte zunächst eine Wirkung durch Gebrauch und Nichtgebrauch der Organe und die Vererbung erworbener Eigenschaften nach Lamarck. Trotz seiner angegriffenen Gesundheit entwickelte er seine Theorie in den Jahren 1837 bis 1842 weiter. 1838 las Darwin das „Essay on the Principle of Population“ von Malthus. Da erkannte Darwin das Prinzip der natürlichen Auslese bei allen Organismen: Jede biologische Art hat die Tendenz zur Vermehrung, die größer ist als die mögliche Vermehrung von Nahrungsmitteln. Dadurch muss es zum Kampf um die Nahrungsquellen, zum Kampf ums Dasein zwischen den Gliedern der Population, zum „struggle for life“ kommen. Darwins origineller Gedanke bestand darin, dass auf einer ersten Stufe eine Überproduktion unterschiedlicher Individuen, „sports“, Spielarten, erfolgt. In einer zweiten Stufe haben die den Lebensbedingungen am besten gewachsenen Tiere und Pflanzen die größte Chance, sich fortzupflanzen. Durch Selektion über große Zeiträume entstehen schließlich neue Arten. Eigene experimentelle Erfahrungen sammelte Darwin auch durch das Studium der Cirripedia (Rankenfüßler) und schrieb ein großes Werk über diese Krebs-Gruppe und ihre Systematik. Die Cirripedien zeigten trotz äußerer großer Unterschiede wie Seepocken (Baelanomorphe), Entenmuscheln (Lepadomorphe) und Wurzelkrebse (Rhizocephalen), z. B. Sacculina, durch das gemeinsame Krebslarvenstadium der Cypris-Larve eine gemeinsame Abstammungsform, von der aus sich die verschiedenen Formen entwickeln. In einem Puppenstadium, der Cyprispuppe, mussten tief greifende Umwandlungen erfolgen.
Erst 1859 fühlte sich Darwin so weit, seine Arbeiten in „Die Entstehung der Arten“ zu publizieren. Die erste Auflage soll am Tage des Erscheinens vergriffen gewesen sein. In seinem Werk vermied er die Fachsprache, so dass seine Theorie auch Nichtbiologen verständlich war. So wurde sie bald überall bekannt. Darwin stellte sich mit seiner Theorie gegen die herrschende Theorie der Artkonstanz, doch lehnte er die Behauptungen seiner Zeit, z. B. in der Naturtheologie, nicht rundweg ab, sondern deutete sie für seine Vorstellungen. Nach seiner Publikation meldete sich Alfred Russel Wallace schriftlich. Er hatte ebenfalls Lyells Ansichten studiert und schrieb, dass auch er von der Veränderlichkeit der Arten überzeugt sei. Doch hatte er auf der Heimreise durch eine Schiffskatastrophe seine frühen Aufzeichnungen über die Forschungsergebnisse verloren. Nach einer neuen Forschungsreise hatte er 1855 „On the Law which has regulated the Introduction of New Species“ veröffentlicht. Im Gegensatz zu Darwin hatte Wallace keinen Mechanismus für eine Artbildung, die Selektion, gefunden. Er wandte sich gegen das Prinzip von der Vererbung erworbener Eigenschaften und gegen die Vererbung von Organveränderungen durch Gebrauch oder Nichtgebrauch. Er lehnte auch Darwins Vorstellung einer Selektion durch reproduktive Isolation zwischen Populationen ab. Darwin glaubte, dass die „Divergenz“, das Auseinandergehen der Individuen, im Verlaufe langer geologischer Epochen zur Artbildung und dadurch einer „Höherentwicklung“ führen könne. Durch die natürliche Auslese werden dabei immer die am stärksten spezialisierten Varietäten und Arten bevorzugt, die dann am wenigsten miteinander konkurrierten.
In seinem Buch von 15 Kapiteln und einer großen Einleitung schildert er den Artenwandel durch natürliche Zuchtwahl im Kampf ums Dasein, die von Landwirten und Gärtnern durch analoge künstliche Zuchtwahl betrieben wird. Zunächst legte Darwin dar, dass die Arten nicht unabhängig voneinander geschaffen wurden, sondern ähnlich den Varietäten von anderen Arten abstammen, wobei als Varietäten zwei Formen bezeichnet wurden, die sehr wenig voneinander differierten. Stärkere Differenzen führten zu Arten und noch stärkere schließlich zu Gattungen. Im dritten und vierten Kapitel stellt Darwin seinen Mechanismus der Artbildung durch das Überleben der Tüchtigsten im Kampf ums Dasein dar, dem „survival of the fittest“ nach Herbert Spencer (1820 – 1903).13 Am Beispiel des Elefanten zeigt Darwin nach der Lehre von Malthus, wie Elefanten, die sich erst nach 30 Jahren, dann aber bis 90 Jahre fortpflanzen, in 750 Jahren 19 Mio. Elefanten bei ungestörter Fortpflanzung hervorbringen. Die Erhaltung vorteilhafter Unterschiede und die Vernichtung nachteiliger nennt Darwin „natürliche Zuchtwahl“. Die Variabilität wird nach Darwin durch die Änderung der Lebensverhältnisse geschaffen. Ohne diese kann die natürliche Zuchtwahl nicht wirken. Die Wirkungen der natürlichen Zuchtwahl auf die Nachkommen gemeinsamer Eltern zeigen sich in der Divergenz der Merkmale. Nur Veränderungen, die nützlich sind, werden erhalten und für die Zuchtwahl verwertet. Das Prinzip der Nützlichkeit kann durch die Divergenz der Merkmale wirksam werden. Die Zuchtwahl häuft sie an. Der Grund, dass es noch nicht entwickelte „niedere Formen“ gibt, liege daran, dass die natürliche Zuchtwahl nicht notwendig einen Fortschritt der Entwicklung bedeute. Auch eine geschlechtliche Zuchtwahl ermöglicht dem Sieger die Fortpflanzung durch Kampf zwischen Individuen eines Geschlechts, i. a. der Männchen, und hängt nicht vom Kampf ums Dasein ab.
Darwins großes Werk war die systematische Erarbeitung des Nachweises der Entstehung neuer Spielarten in der Natur, die letztlich analog auf experimenteller Basis von Züchtungen bei Tieren und Pflanzen gewonnen werden können, die also von anderen abstammen: Deszendenztheorie. Als Ursache in der Natur erkennt er die Selektionsbedingungen einer sich ändernden Umwelt durch natürliche Zuchtwahl auf zufälliger Basis: Selektionstheorie. Die Erklärung der Variation erforderte Kenntnis der genetischen Gesetze und der molekularbiologischen Prozesse, die Darwin ohne die Erkenntnisse Mendels und bei dem Stand der damaligen Forschung verborgen bleiben mussten. Aber der Gedanke einer „zufälligen" Variation der Spielarten war grundgelegt.
Darwin diskutiert in seinem Buch in den folgenden 11 Kapiteln die Möglichkeiten einer weiter führenden Höherentwicklung auf der Basis seiner gefundenen Prinzipien der Variabilität und Selektion. Dabei nahm er an, dass sich in den verschiedenen biologischen und paläontologischen Disziplinen die erforderlichen Beweise mit fortschreitender Forschung, die „missing links“, auch in der Erdgeschichte finden werden.
Heute muss im Lichte der genetischen Erkenntnisse festgestellt werden, dass sich der experimentell prüfbare Artenwandel im Bereich der sog. Mikroevolution oder intraspezifischen Evolution (nach Bernhard Rensch) bewegt. Dabei handelt es sich genetisch um eine Umkonstruktion des artspezifischen Erbmaterials und damit vorgegebener Organisationsmale, Strukturen und Baupläne, einer genetischen Veränderung in der Horizontalen des Genbestandes. Die Mikroevolution erweist sich bei plötzlichen Umweltveränderungen als Überlebensstrategie, weil die angepassten Spielarten oder Varianten eines Typs bereits vor der Umwälzung vorlagen. Solche grundlegenden unterschiedlichen Ausprägungen eines Konstruktionstyps zeigen die flugunfähigen Insekten einer Insel, die Helligkeitsvarianten des Birkenspanners, die Farbmutanten des Wellensittichs, die Blattmutanten der Erbsen, aber auch die weiterführenden Typvariationen, z. B. der Rosengewächse und Orchideen, bei Tieren die hunde-, pferde- oder katzenartigen, bei Bakterien die Myxobakterien oder Aktinomyceten. Sie sind nicht nur durch morphologische, anatomische, physiologische Ähnlichkeiten miteinander verbunden, sondern auch durch die Möglichkeit einer Bastardisierung mit wenigstens einem Beginn einer Embryonalentwicklung oder bei Bakterien durch eine sehr ähnliche Genfolge auf dem Chromosom. Die Entstehung der Varianten, Darwins „sports“, fanden erst 1907 in der Entdeckung der Mutationen eine genetische Basis, die in den 20er Jahren des vergangenen Jahrhunderts durch Strahlen und in den 30er Jahren durch Chemikalien ausgelöste, „induzierte“ Mutation erweitert werden konnten: Nobelpreisträger Werner Arber unterscheidet heute drei experimentell prüfbare Strategien einer evolutionären (d. h. mikroevolutiven) Entwicklung: Punktmutationen, bei der der Austausch eines einzigen Nukleotidpaares der DNA eine Veränderung der Eigenschaften bewirken kann, ferner DNA-Umstrukturierungen und den DNA-Erwerb, z. B. extrachromosomale DNA als Plasmide bei Bakterien. Gene können als „Generatoren“ der genetischen Variation auftreten, wobei andere Gene die genetische Plastizität beschränken. Eine gezielte Richtung ohne natürliche Selektion zu geben, vermag nur der Experimentator im Labor durch ausgeklügelte Techniken mit entsprechendem planerischem und materiellem Versuchsaufwand in der DNA-Synthese in vitro mit ausgeklügelter Sequenzvariation.
3. Über die Entwicklung von Darwins Werk zur modernen Evolutionstheorie
Darwin baute auf seiner Theorie der Entstehung der Arten die Hypothese einer „Höherentwicklung“, einer Evolutionstheorie auf, und schildert in den folgenden 11 Kapiteln seines Werkes die Nachweise dafür. Er verließ damit den experimentellen Bereich der vollständigen Induktion und extrapolierte die mikroevolutionär gesicherten Vorgänge auf nicht experimentell prüfbare Vorgänge der „transspezifischen Evolution“ (Bernhard Rensch) oder Makroevolution, einer Neukonstruktion von Erbmaterial, der Bildung qualitativ vollständig neuer Gene. Als Grenzfall kann auch die Entstehung des Lebendigen, die chemisch-biologische Evolution, eine Entstehung des Lebens aus anorganischen Stoffen betrachtet werden. Die Begriffe Mikro- und Makroevolution sind in der Evolutionsbiologie schon lange im Gebrauch, z. B. von Bernhard Rensch (1900 – 1990) als „intra- und trans-spezifische Evolution“ und wurden bereits 1927 von Philiptschenko eingeführt.14 Makroevolutionäre Übergänge zeichnen sich durch die Notwendigkeit zahlreicher unabhängiger Vorgänge aus, die ähnlich einem Puzzlespiel genau zusammenpassen. Sie werden durch latente oder entwicklungsbiologische Potentiale nur vorgetäuscht, wie bei Genduplikationen, Neukombinationen von Genen (Exon shuffling), durch Mutanten von Hoxgenen u. a. Sie lassen sich stets auf mikroevolutionäre Vorgänge zurückführen. Von Darwins Anhängern wurden ganze Reihen von Erkenntnissen zusammengetragen und hypothetisch nach der generalisierenden Induktion in Entwicklungszusammenhänge gebracht. Dazu wurden verschiedene Disziplinen der Biologie/Paläontologie für Beweise herangezogen, in der Hoffung, die noch vorhandenen Lücken durch weitere Forschung zu füllen.
3.1. Die Belege
(1) Die Paläontologie. Die Fossilien sollen den tatsächlichen Ablauf der Stammesgeschichte dokumentieren. Die Höherentwicklung sollte nach Darwin allmählich, gradualistisch, verlaufen. Der Geologe Otto Heinrich Schindewolf (1896 – 1971) aber vertrat einen sprunghaften, punktuellen Verlauf, wie aus seiner Formulierung 1948 hervorgeht: „Der erste Vogel kroch aus dem Reptilienei“. Nach den sprunghaft auftretenden Fossilien unbekannter Arten ließ sich eine gradualistische Stammesgeschichte kaum halten. Daher wird heute eine punktuelle Phylogenie vertreten: Phasen der Ruhe wechseln mit solchen rascher Veränderung. Ein phylogenetisches Forschungsprogramm entstand. Eine phylogenetische Entwicklungsreihe zeigte schon im 19. Jahrhundert Kowalevski (1842 – 1883) mit der Abstammungsreihe des Pferdes vom Eohippus (Eozän) mit vier Zehen über Orohippus (Eozän) und Hipparion (Pliozän) bis zum heutigen Unpaarhufer, die mit einer Adaptation an die Grassteppe gedeutet werden kann.15 Auch die Mikrobiologie kann ein „Paradepferd“ in einer Stoffwechselmutante aufweisen (siehe 3.4).
(2) Die Vergleichende Anatomie und Morphologie lieferte Darwin die meisten Argumente. Richard Owen (1804 – 1892) brachte den Begriff des „homologen Organs“, das bei verschiedenen Organismen das gleiche Bauprinzip zeigt, aber verschiedene Funktionen besitzt, die auf evolutionärem Wege abgewandelt worden sein sollen. Auch die Vorderextremitäten der Wirbeltiere weisen Abwandlungen in verschiedener Funktion auf: Flügel bei Fledermäusen oder Grabbeine beim Maulwurf. Diesen homologen Organen stehen analoge Organe gegenüber, in gleicher Funktion, aber bei offensichtlich fehlender gemeinsamer Abstammung: die Grabbeine bei Maulwurf und Maulwurfsgrille. Alle homologen Merkmale versuchte Darwin durch die Wirkung der natürlichen Auslese zu erklären. So sollten auch nach der Reichert-Gauppschen Theorie die Gehörknöchelchen Hammer und Amboss sich von den Kiefergelenkknochen Artikulare und Quadratum der Reptilien herleiten, wodurch das Gelenk zwischen Hammer und Amboss als „primäres Kiefergelenk“ gewertet wurde, so dass bei Säugetieren die Bildung des Squamoso-Dentale-Gelenks aus anderen Kieferknochen der Reptilien zum sekundären Kiefergelenk wurde. Aufgrund neuer Fossilfunde muss heute jedoch davon ausgegangen werden, dass sich die Gehörknöchelchen mindestens zweimal unabhängig voneinander entwickelt haben.16 So ist es nicht mehr möglich, aus der Homologie sicher auf eine gemeinsame Abstammung zu schließen. Lebewesen, die Merkmale verschiedener Gruppen zeigen, werden als Zwischenformen bezeichnet. Dazu werden Quastenflosser (Amphibien und Fische), Lungenfische (Fische und Amphibien), Ichthyostega (Uramphib), Seymouria (Urreptil), Archaeopteryx (Urvogel), Synapsiden (Ursäuger) und das Schnabeltier als eierlegendes Säugetier mit Kloake (wie bei Vögeln und Reptilien) angeführt.
(3) Die Embryologie wurde als Werkzeug zur Erkennung einer Homologie benutzt. Schon vor Darwin wurde 1821 von Johann Friedrich Meckel (1781 – 1833) behauptet, ein höheres Tier durchlaufe in seiner Entwicklung die unter ihm stehenden bleibenden Stufen.17 Daraus folgt auch, dass die Entwicklungsgesetze der Ontogenie und Phylogenie die gleichen seien. Dies führte 1866 Ernst Haeckel (1834 – 1919) zum sogen. Biogenetischen Grundgesetz: „Die Ontogenie ist eine kurze Rekapitulation der Phylogenie“. Danach müssten die Organismen in ihren Entwicklungsstadien ihren Vorfahren ähneln, weil sie aus ihnen entstanden sind. Dies bedeutet aber, dass die Evolution nicht zufällig ist, da Ontogenie und Phylogenie den gleichen Gesetzen gehorchen. Die Entwicklung des Amphioxus wurde zur typischen Ontogenese der Wirbeltiere. Die Gastraea-Theorie Haeckels brauchte eine Anwendung des biogenetischen Grundgesetzes. Dieses schien sich in der Gastraea-Form zu bieten. Sie sei eine Urform aller Metazoen, womit der Ursprung aller Metazoen nachgewiesen schien. Heute gilt, dass die Ontogenie der Gastraea-Theorie in vielen Fällen nicht folgt und von der Phylogenie unabhängig verstehbar ist, z. B. konnte Blechschmidt18 zeigen, dass die Ontogenie des Menschen unabhängig von der postulierten Phylogenie für sich verständlich ist. Das Biogenetische Grundgesetz gilt allenfalls als Regel. Nichtsdestoweniger hat es zu Forschungsprogrammen geführt.
(4) Die Systematik. Vor Darwin war das natürliche System Ausdruck eines ideellen Ganzen. Darwin brachte es als genealogisches System auf materialistischer Grundlage. Julius Wilhelm Albert Wiegand (1821 – 1886), Vertreter des Prinzips der Arten, erkannte 1872 das genealogische Prinzip gezwungenermaßen an.19 Die erbliche Einheit ließ aber keine Aussage über den Artenwandel zu.
Der „Typus“ vor Darwin wurde durch Darwin zum „Vorfahren“, von der „figürlichen Verwandtschaft“ führte der Weg zur „Blutsverwandtschaft“, d. h. Abstammungsverwandtschaft. Diese gemeinsame Verwandtschaft wurde folgerichtig dann auch auf den Menschen ausgedehnt.20 Dass des Menschen Geist und sein sittliches Gewissen materielle Naturprodukte sein sollen, stieß auf Ablehnung. Daher sprach Darwin vorsichtig von „Light will be thrown on the origin of man and his history“.21 Ernst Haeckel entfaltete Darwins Lehre in dem dreibändigen Werk „Ein natürliches System der Organismen aufgrund ihrer Stammesgeschichte in Systematischer Phylogenie“. Heute sind 16S- und 18S-Analysen der ribosomalen Ribonukleinsäuren und deren Gen-Sequenzen der allen Organismen gemeinsamen zellulären Proteinsynthese auf DNA-Niveau möglich. Sie begründen Cladogramme, Stammbäume, auf molekularer Grundlage, die sie exakt erscheinen lassen. Obwohl nicht wenige Ausreißer zu bedenken sind, ist die molekularbiologische Basis für die Verwandtschaftsbestimmung in einem natürlichen System heute unentbehrlich. Es bleibt die Frage, ob diese den tatsächlichen Verlauf einer postulierten Stammesgeschichte widerspiegeln. Es ist unbestreitbar, dass die 16S-rRNA-Analysen (für Prokaryonten) und 18S-rRNA-Analysen (für Eukaryonten) Forschungsprogramme enorm befruchtet haben.
(5) Die Biogeographie. Sie beruht auf Darwins Beobachtung, dass sich nahe verwandte Arten geographisch ersetzen können. Die Faunen von Inselgruppen sind meist denen des benachbarten Kontinents ähnlich. Dies lässt auf eine Abstammungsverwandtschaft schließen, wie dies Darwin für die Fauna der Galapagos-Inseln getan hat. Hier wurde klar, dass die Faunenverwandtschaft weniger durch die Umwelteinflüsse als durch die gemeinsame Abstammung bestimmt wird. So entspricht die Vegetation Englands der Europas. Viele Arten haben, wie Darwin fand, eine große Wanderungs- und Ausbreitungsfreudigkeit. Der Mensch ist hier ein extremes Beispiel. Arten, die sich in bereits bewohnten Gebieten ansiedeln wollen, müssen u. a. mit den Ureinwohnern den Kampf ums Dasein bestehen.
An die Verdienste des Forschers Wallace um die Biogeographie sei erinnert (Kap. 1). Darwins biographische Untersuchungen waren für seine Speziation, die Artbildung, wichtig. Hierbei spielte die Migration mit geographischer Isolation eine große Rolle, die später als wichtiger „Evolutionsfaktor“ gewertet wurde. Gleichzeitiges „sympathisches“ Zusammenleben erklärte Darwin durch vorausgegangene klimatische und geologische Isolation. Darwin gestand auch ein, dass Arten durch verhaltensbiologische oder ökologische Isolation entstehen könnten. Er übertrug dies auf die menschliche Rassenbildung. 1870 stand die Selektionstheorie Darwins als Alternative Rudolph Wagners (1805 – 1864) Separationstheorie gegenüber und wurde auf die Rassenbildung des Menschen angewendet.22
Heute ist festzustellen, dass zahlreiche Forschungsarbeiten mit biographischen Themen aus Darwins Pionierarbeit hervorgegangen sind, die ihn zum Begründer der kausalen Biogeographie werden ließen.
(6) Ökologie. Nach Darwin war die Sexualität für die Evolution notwendig; denn nach seiner Überzeugung war die Selbstbefruchtung bei Pflanzen in der Natur nicht der Normalfall. Er entdeckte die wechselseitige Anpassung von Blüten und den sie besuchenden Insekten, die Koadaptation. Darwin sah die Nicht-Selbstfruchtbarkeit als Naturgesetz an. Hermaphrodite Pflanzen besitzen auch sexuelle Fortpflanzung, was Darwins Selektionstheorie abschwächte.
1862 entdeckte Henry Walter Bates (1825 – 1892) die Mimikry, die Nachahmung wehrhafter durch nichtwehrhafte, in Körpergestalt und Färbung, z. B. fressbare und giftige Schmetterlinge.23 Bates konnte solche Anpassungen experimentell zeigen. Auf dem Gebiet entstanden ebenfalls große Forschungsprogramme unter Anwendung der Selektionstheorie.

3.2 Die moderne Evolutionstheorie (Abb. 2)
Durch das Bekanntwerden der Ergebnisse der Genetik, wie der Rekombination, die mit sexueller Fortpflanzung einhergeht, der Mutation als Basis der Variabilität der Konstanz der Arten als Voraussetzung von Vererbung und Systematik, konnte die Evolutionstheorie schrittweise weiter entwickelt werden. Daneben können der Gentransfer, die Übertragung von Genen, z. B. bei Bakterien (1944) durch Transformation (Oswald Theodor Avery, 1877 – 1955), und die Gendrift (zufällige Mutationen, die selbst keine Änderung des Genbestandes bewirken (Neutralmutationen), weil sie sich zunächst nicht bemerkbar machen, aber später spontanen Einfluss z. B. auf benachbarte Gene ausüben können), genannt werden. Es ist der enge Bereich zwischen Veränderlichkeit und der Konstanz der Arten, der das „Rohmaterial der Evolution“ in den verschiedenen Varianten schafft. Die Mutationen werden i. A. als „spontan“ bezeichnet, wenn die Ursache nicht erkennbar ist. Fehlpaarungen, zahlreiche von außen auf den Organismus wirkende Mutagene, wie Radioaktivität oder ultraviolette Strahlung, verschiedene chemische Verbindungen, die auch Krebsgifte sein können, aber auch Hitze und Säuren, und „innere Mutagene“ wie bestimmte Stoffwechselprodukte, sind bekannt. Die Überproduktion, mitunter regelrechte Populationswellen, ermöglichten der Selektion die „richtunggebende Kraft“ der Evolution. Eine Fülle von Selektionsfaktoren, wie die Isolation, z. B. die geographische (Gebirge), ökologische und sexuelle Isolation, das Nahrungsangebot, die Atmosphäre, die z. B. bei bakteriellen Spezialisten Schwefelwasserstoff benötigt, und Sauerstoff, der bei anaeroben Bakterien sogar tödlich wirken kann, Lichtverhältnisse bei Photosynthetisierern, Klimaveränderungen, Temperaturen und Umweltgifte wirken bei der Auslese der „survival of the fittest“. Bei der Erzeugung der Vielfalt der Varianten, der Mutanten, wirkt der Zufall, d. h. völlig ungerichtet tritt an der DNA im Genom eine Mutation auf, z. B. in der chemischen Veränderung eines Nukleotids, ein DNA-Kettenbruch mit Neuverkettung, Bruchstückaustausch, Reversionen der Bruchstücke oder „jumping genes“, die durch Integration oder Ausgliedern Gene ein- oder ausschalten können. Die weitere Ausprägung zur Erbfestigkeit erfolgt naturgesetzlich! Aus der großen Vielfalt werden Mutanten eines „Pools“ durch die Selektionsbedingungen der Umwelt herausgelesen, d. h. für die Selektionsbedingungen liegen schon die verschiedensten Mutanten für die nachfolgende Selektion bereit. Es liegt keine zielgerichtete Planung vor wie beim Lamarckismus, bei dem sich z. B. die Giraffe nach dem Laub der höher gelegenen Äste reckt, wodurch die Hälse allmählich länger werden. Dagegen werden nach der Evolutionstheorie die rein zufällig entstandenen am besten angepassten Mutanten von der Umwelt nur ausgewählt. Nur das Selektionsprinzip wirkt als „richtunggebende Kraft“ (Abb. 3). Da zur Zeit Darwins Mendels Erbgesetze nicht bekannt geworden waren, hatte man lamarckistische Mechanismen zugelassen, die die Entstehung der Variabilität erklären sollten. Sie dienten auch Darwin zur Ergänzung der Selektionstheorie, nach der die Umwelt die phylogenetische Entwicklung über einen lamarckistischen Mechanismus ermöglicht. Die stammesgeschichtliche Entwicklung über zielgerichtete innere Faktoren war populär.
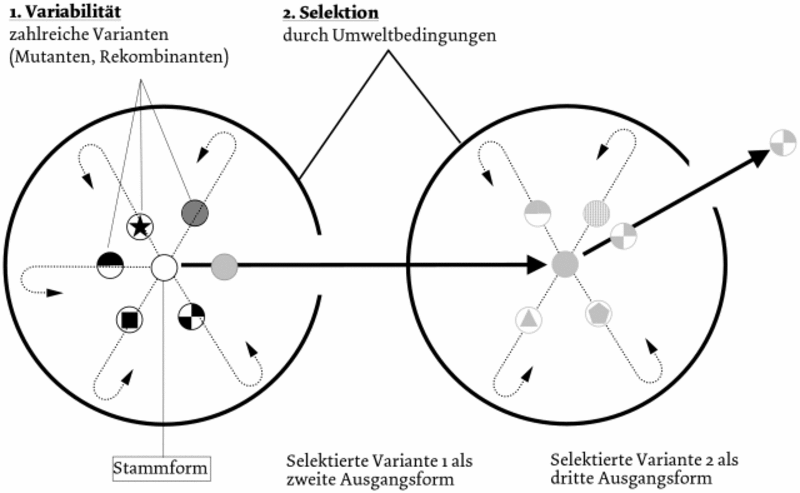
Die moderne Evolutionstheorie erscheint zunächst als eine Beschreibung einer möglichen Entwicklung, die als solche angesichts anderer in der Biologie bekannter Entwicklungsvorgänge kaum bezweifelt werden kann, wie die Ontogenese, selbst die Wachstumsvorgänge und die Entstehung von Lebensgemeinschaften. Die Schwäche ist die Annahme eines sehr einfachen Mechanismus, nämlich des Zufalls, der die Entstehung höchst komplexer Einrichtungen ermöglichen soll, deren Konstruktion ohne Abarbeiten eines vorgegebenen Planes der Erfahrung widerspricht. Nach Carl Wilhelm von Naegelis (1817 – 1891) Idioplasmatheorie (1884) besitzt jedes Individuum eine innere Kraft, die unter den äußeren Bedingungen, unter denen es aufwächst, prägend wirkt.24 Naegeli verglich diesen Prozess mit einem Kristallisationsvorgang. Er lehnte sowohl die Selektionstheorie als auch die gemeinsame Abstammung aller Organismen ab und postulierte eine Unzahl phylogenetischer Stämme ohne genetische Verwandtschaft. August Weismann (1834 – 1914) prüfte die Selektion experimentell, indem er Raupen auf unterschiedlich gefärbten Untergrund aufsetzte und sie dann Räubern aussetzte, welche die Raupen je nach Untergrundfarbe verschieden wahrnahmen.
Die Selektion durch zahlreiche Umweltfaktoren in Verbindung mit den wieder entdeckten Vererbungsgesetzen und der Variation durch Mutation und Rekombination ohne die Vererbung erworbener Eigenschaften prägte die Zeit des Neodarwinismus. In der Zeit der Synthethischen Evolutionstheorie wurden Mikro- und Makroevolution, Phänotyp und Genotyp definiert. Schließlich kam die Endosymbiontentheorie, die Mitochondrien und Chloroplasten als eingewanderte umgewandelte Prokaryonten sieht, auf. Wichtiger Forschungsgegenstand sind heute molekulare Stammbäume und Membranforschung.
3.3 Diskrepanzen zwischen Evolutionslehre und Forschungsergebnissen
Die Fortschritte der Wissenschaft zeigen gerade in letzter Zeit erstaunliche Erkenntnisse, die Phantasie und Kreativität herausfordern.
3.3.1 Rätsel für die Evolutionstheorie
Es sollen ein paar Beispiele angeführt werden, die mit der heutigen Makroevolution nicht harmonisierbar sind. Der Mangel an Übergängen „missing links“ bei höheren Taxa ließ sich durch die zahlreichen, aber diese nicht beweisenden Fossilien nicht ausgleichen. Nur vollendete Organismen mit vollständigen Einrichtungen sind sowohl bei rezenten Arten als auch Fossilien nachzuweisen. Der Angriff der Selektion an unvollendeten Einrichtungen ist rätselhaft. Ein schubweiser Evolutionsverlauf punktueller Natur wäre zu postulieren. In gänzlich verschiedenen Organismen wurden merkwürdigerweise ähnliche Gene gefunden. Dies spricht für eine voneinander unabhängige Entstehung. Einrichtungen für die wichtige Energiegewinnung, wie Photosynthese und Oxidationen organischer oder anorganischer Stoffe oder die Kohlendioxidassimilation, treten mehrfach unabhängig voneinander auf, nur die Stickstoff-Fixierung scheint auf einen einmaligen Entstehungsvorgang zurückzuführen sein. Man kann hier von einer funktionalen Konvergenz sprechen. Unter Konvergenz versteht man bauplanähnliche Strukturen, die mehrmals unabhängig voneinander entstanden sind und keine Basis für eine gemeinsame Abstammung bieten: Die Torpedoform von Haien, Fischsauriern, Delphinen und Pinguinen oder die Fangbeine der Netzflügler und Heuschrecken (Mantis). Ein neuer Fall konvergenter Evolution, der bisher als Beleg (Schlüsselmerkmal) für Makroevolution gehalten wurde, sind die schon erwähnten Gehörknöchelchen (s. Kap. 3)25 Hammer, Amboss und Steigbügel. Bei Säugern sollen sie vor der Abspaltung der Kloakentiere aus dem primären Kiefergelenk mit Angulare, Artikulare und Praeartikulare, letztere verschmolzen zum Hammer, hervorgegangen sein, so dass als Unterkieferknochen nur das Dentale übrig blieb, das das sekundäre Kiefergelenk mit dem Squamosum bildet. Beuteltiere und Placentalier sollen 215 – 225 Mio. Jahre alt sein und die Kloakentiere entstanden 100 Mio. Jahre später. Nun wurde ein Kloakentier Teinolophos trusleri, 115 Mio. Jahre alt, fossil gefunden. Dieses besaß aber außer dem Dentale noch weitere Unterkieferknochen. Bisher hatte man die Gehörknöchelchen aus dem reptilischen Kiefergelenk für alle Säugetiere vor der Abspaltung gemeinsam entstanden gedacht. Doch nun verlieren die komplizierten Gehörknöchelchen den Charakter als typisch für die ganze Säugetiergruppe, die makroevolutionär einmal entstanden seien. Die zweimalige unabhängige Entstehung aber bedeutet konvergente Evolution und stellt das Hervorgehen der Gehörknöchelchen aus den Unterkieferknochen der Reptilien in Frage und erschwert die Theorie der gemeinsamen Abstammung. Erstaunlich ist, dass die ältesten Cyanobakterien mit ihrer komplizierten Photosynthese von einem Alter bis 3,5 Mrd. Jahren zu 30% mit heute lebenden Arten identifiziert werden können. In Bernstein eingeschlossene Organismen von einem geschätzten Alter von 30 Mio. Jahren und älter lassen sich bis in feinste Strukturen mit heute lebenden Organismen identifizieren. In all diesen Fällen wurden keine wesentlichen Veränderungen gefunden. Sogar Mikroorganismen (Bacillus circulans) in Gesteinen in 140 Mio. Jahre altem Zechstein z.B. wurden lebensfähig gefunden und konnte mit heutigen Stämmen identifiziert werden.26 Infektionen wurden von zahlreichen Arbeitsgruppen auf diesem Gebiet ausgeschlossen. Stammbäume lassen sich aufgrund der Gensequenzen der 16S rRNA und der 18S rRNA konstruieren. Sie stimmen aber nur für ausgewählte Organismen einer Gruppe. Stammbäume der Cytochrome und bestimmter Enzyme führen bei Überlagerung zu netzartigen Gebilden und keineswegs zu einer deckungsgleichen Übereinstimmung. Anstelle der Haeckelschen Stammbäume findet man ausgedehnte Netzwerke von Artengruppen. Als Urahne kommt keine einzelne Zelle in Frage, sondern eine Zellpopulation. Die Tatsache, dass seit 600 Mio. Jahren keine neuen Organismenstämme mit neuen Konstruktionsplänen nachzuweisen sind, wird als „Rätsel der Evolution“ bezeichnet. In relativ kurzer Zeit entstanden diese neuen Baupläne, „Kambrische Explosion“ genannt. Explosionsartig sind auch im frühen Kambrium komplexe Gehirne aufgetreten und haben seitdem ihre Struktur kaum verändert. Der Genetische Code, bei Gebrauch mit minimalen Kopierfehlern, erwies sich unter 1 Mio. Fällen als einmalig nach Computerberechnungen und gilt als „nicht durch Zufall entstanden“.27 Auch die Entstehung der Sexualität ist nicht durch Selektion erklärbar. Zurzeit gibt es dafür kein Modell, das geeignet wäre, eine einheitliche Erklärung nach der Evolutionstheorie zu liefern.28
3.3.2 Komplexe Einrichtungen (siehe auch Kap. 6)
Während der Weg von komplexen Einrichtungen zu einfachen durch Defektmutationen oder Genverluste bei Spezialisierungen leicht verfolgt werden kann, bleibt der umgekehrte Weg einer Höherentwicklung rätselhaft. Wie erwähnt, konnte keine überzeugende Entstehung von qualitativ neuen biologischen Strukturen beobachtet werden. Ein äußerst komplexes Objekt ist der ca. 24 nm große Geißelmotor, der Bakterien zur Fortbewegung dient. Er ähnelt einem Elektromotor mit Stator und Rotor.29 Als Antrieb wird ein Ionenstrom angenommen. Die Umdrehungsgeschwindigkeiten pro Minute betragen beim Protonenstrom 18.000 und beim Natriumionenstrom 48.000. Diese Umdrehungsgeschwindigkeiten bewegen sich in der Größenordnung moderner Ultrazentrifugen. Für die komplette Funktion von Geißelmotor und gesteuerter Fortbewegung sind ca. 60 Gene erforderlich, die mehr als 2% des Bakteriengenoms einnehmen. Zur Funktion müssen alle Gene intakt sein. Wie könnte an einem Entwicklungsteilstadium die Selektion angreifen? Zudem würde ein funktionsloses Übergangsstadium als unökonomische Belastung für die Zelle wieder degenerieren. Wie könnten die zahlreichen Mutationen in den Genen die erforderliche Qualität für die Bauteile mit ihren Feinstrukturen und Passformen des Geißelmotors erhalten?
Der Zoologe Adolf Portmann (1897 – 1982) wies daraufhin, dass auch soziale Verhaltensweisen bei Tieren, Gestaltungen und Strukturen über bloße Erhaltungsstrukturen hinausgehen.30 In ihrer Entstehung könnten Mutanten nur die Organausformung und den Darstellungswert erhöhen. Auch bei Pflanzen seien die Ursprünge der Blütengestaltung und Formung auffälliger Früchte rätselhaft, da die Selektion erst am Vorhandenen und Vollendeten angreifen könne. Überraschend war, dass auch Bakterien soziobiologisches Verhalten aufweisen, wie es sonst nur bei höheren Organismen bekannt ist, z. B. bei Myxobakterien. Von den Prokaryonten Archaeen, die wegen ihrer Spezialisierung auf extreme Umweltverhältnisse, wie Vermehrung bei einer Spezies bei 113 °C, für primitiv gehalten wurden, sagte ein Evolutionsbiologe, erstaunt über seine Entdeckungen, „die sind ja so kompliziert wie wir“.
3.3.3 Über die Arbeiten von Grundlagen zur Entstehung des Lebens
Die beiden Stoffgruppen Nukleinsäuren und Proteine gelten heute als Grundlage des Lebens. Die Evolutionstheorie nimmt an, dass die Kondensationsreaktionen von Nukleotiden und Aminosäuren spontan in Ursuppen möglich seien. Der Polymerenchemiker Bruno Vollmert hat in seinen Arbeiten klar nachweisen können, dass alle publizierten Versuche zur Kondensation von Nukleotiden und Aminosäuren für das Problem der Evolution auf Molekülebene irrelevant sind, da sie nur mit reinen Monomeren und nicht unter Ursuppenbedingungen entstehen können. Polykondensationen in Ursuppen und den darin gelösten Stoffgruppen „sind ebenso wenig möglich, wie die Konstruktion eines Perpetuum mobile“31. Schon Spuren von monofunktionellen Verbindungen bewirken stets einen Kettenabbruch. Heterogenkatalysen sind ohne Bedeutung.
Stanley Miller ging der Frage nach, wie die Vorstufen der Makromoleküle, Nukleotide und Aminosäuren, in einem Urmeer mit einer Uratmosphäre aus Methan, Ammoniak, Wasserstoff und Wasser unter elektrischen Entladungen bei höheren Temperaturen entstehen könnten. Der Chemiker weiß, dass unter diesen Bedingungen hoch reaktive Radikale entstehen, die in einem breiten Spektrum miteinander reagieren. Er fand Verbindungen, die zum Teil in Organismen vorkommen, aber andere wichtige Verbindungen fehlten. Organische Verbindungen können unter ähnlichen Bedingungen auf Planeten und in Kometenmaterial vorkommen. Jedoch bringen solche Ergebnisse die Frage der Entstehung des Lebens keinen wesentlichen Schritt einer Lösung näher.
Nach der Evolutionstheorie wird das Nebeneinander von Organismen und Einrichtungen verschiedenen Komplexheitsgrades – der als das Ergebnis einer Höherentwicklung verstanden wird, und die Differenzierung durch Zufall, also ohne Plan, jedoch nach dem Nobelpreisträger Manfred Eigen „eingeschränkt auf Naturgesetzlichkeit“, die nicht näher bekannt ist – mit anschließender Selektion der „durchsetzungsbefähigten“ Moleküle oder Organismen erklärt.32 Dabei sind die besser angepassten Formen (Mutanten) durch eine statistische Fluktuation möglich. „Zielscheibe der Selektion ist die gesamte Verteilung… die bestangepasste Sequenz" der Nukleotidkette. Die „Realisierung der Schöpfung, die Evolution ist“, geschieht nach Naturgesetzlichkeit einer letztlich unbekannten Triebfeder, die sich wissenschaftlichem Zugriff entzieht.
3.4 Evolution bei Bakterien unter unseren Augen
So bezeichnete ein Wissenschaftler die bei Bakterien im Labor zu beobachtende Evolution. In verhältnismäßig einfachen Versuchen lässt sich die Resistenzbildung bei Bakterien verfolgen. Sie ist für die Medizin ein Problem bei der Behandlung von Infektionskrankheiten und für die Bakterien eine Frage der Überlebensstrategie.33
Bei der Resistenz gegen Spektinomycin lässt sich z. B. an der Bindungsstelle des Antibiotikums am S5-Protein an der kleinen Untereinheit der Ribosomen ein Aminosäureaustausch von Serin gegen Prolin beobachten. Die dadurch verursachte Änderung der Raumstruktur verhindert die Bindung des Spektinomycins am Wirkungsort.
Bei dem Antibiotikum Chloramphenicol (Chloromycetin) kann das Antibiotikum durch eine Acetylierung des Moleküls unwirksam gemacht werden. Hierzu wird das zuständige Gen auf dem Bakterienchromosom entsprechend der Konzentration des Antibiotikums reversibel vervielfältigt (Genduplikation/Genamplikation), so dass die Acetylierung entsprechend aktiviert wird.
Das Antibiotikum Penicillin aus dem Pilz Penicillium wurde chemisch abgewandelt. Es schädigt die Biosynthese der Bakterienzellwand. Die Bakterien produzieren die Penicillinase, eine Lactamase, die das Molekül Penicillin zerstört und damit zur Resistenz führt. Ein Komplexitätszuwachs ist hierbei nicht zu beobachten. Die Veränderungen liegen im Bereich der Mikroevolution. Dies gilt auch für das „Paradepferd der mikrobiellen Evolution“. Einem Stamm von Escherichia coli wurde das Abbau-Gen für Lactose aus dem Bakterienchromosom herausgeschnitten, so dass es sich auf Lactose als Kohlenstoffquelle nicht mehr vermehren konnte. Nach längerer Zeit jedoch ließ sich mit Spezialmedien doch wieder eine Vermehrung mit Lactose feststellen. Die genaue Untersuchung ergab, dass ein anderes Gen an einer ganz anderen Stelle eingesprungen und mikroevolutionär für die Lactose-Metabolisierung aktiviert worden war.
3.5 Die Information als grundlegende Komponente des Lebendigen
Es ist unbestritten, dass die Information eine grundlegende Komponente des Lebendigen ist. Dennoch werden die Erkenntnisse der modernen Informatik in den Lehrbüchern kaum berücksichtigt. Norbert Wiener sagte: „Information ist Information, weder Materie noch Energie“. Ähnlich drückte sich der Wissenschaftler J. Peil trotz atheistischem Umfeld in der ehemaligen DDR aus: „Information ist kein physikalisches oder chemisches Prinzip wie Energie und Substanz, auch wenn sie ihrer Träger bedarf“.34 Die Übertragung der Information geht von der Informationsquelle, dem nach Werner Gitt mit Intelligenz und Willen begabten Sender, aus und mündet in der Informationssenke, dem gleichfalls mit Intelligenz und Willen ausgestatteten Empfänger. Der Code, der die Bedeutung der in Zeichen übertragenen Botschaft verschlüsselt enthält, muss dem Empfänger bekannt sein. Die Information ist an materielle oder energetische Träger gebunden, z. B. Bleistiftspuren auf dem Papier oder Radiowellen. Der Informatiker, Mathematiker und Ingenieur Werner Gitt sagt zur Makroevolution aus: Da der Mensch in der Lage sei, Information zu schaffen, könne diese nicht von einem rein materiellen Körper stammen. Daraus folgt, dass er über eine nicht-materielle Komponente verfügen müsse. Dem entspricht nach Gitt auch, dass die Behauptung des wissenschaftlichen Materialismus, das Universum sei aus der Materie hervorgegangen, falsch ist. Da die Information nicht von der Materie stammen kann, ist ein immaterieller Sender erforderlich, der nur an seinen Wirkungen in den Naturobjekten erkennbar ist. Ferner, da die Größe „Information“ eine geistige Größe ist, kommen für das Leben keine rein materiellen Prozesse in Frage. So haben alle Versuche wie Computer-Simulation für eine Höherentwicklung und Darwin-Experimente, z. B. mit Bakteriophagen und deren Bestandteilen (Nucleinsäuren und Enzyme als Biokatalysatoren), keine entscheidenden Erkenntnisfortschritte gebracht, so erstaunlich die Anpassungsreaktionen bei entsprechenden Milieuvorgaben auch sein mochten.
Nichtsdestoweniger hält Manfred Eigen an der Hypothese fest, Information entstehe aus Nicht-Information.35 Es sei aber kein Naturgesetz, kein Prozess und kein Ablauf in der materiellen Welt bekannt, nach der in der Materie Information von selbst entstehe, stellt Werner Gitt fest. Zu dieser Erkenntnis kam auch die „7. Internationale Konferenz über die Ursprünge des Lebens“ mit dem 4. Kongress der „International Society for the Study of the Origin of Life (ISSOL)“ in Mainz. K. Dose präzisierte: Ein weiteres Rätsel bleibt die Frage nach dem Ursprung der biologischen Information, also der Information in unseren Genen“. Daraus folgt, dass die Makroevolution, die für die Höherentwicklung die Entstehung qualitativ neuer Gene nach der Informationslehre unmöglich ist und immer nur eine Theorie aus vielen unbeweisbaren Hypothesen neben Tatsachen bleiben und als solche weitere Forschungsprogramme initiieren wird, wie die Hypothese der Atome und Moleküle für die kinetische Gastheorie Forschungsprogramme für die Gasgesetze geliefert hat.
Nach Wolfgang Kuhn36 und anderen, die die Evolutionstheorie bejahen, lässt sich das Leben nicht allein aus Materie und Energie, sondern nur unter Hinzunahme des geistigen Prinzips Information charakterisieren. Die Herkunft von Materie und Energie ist dabei sowieso ein ungelöstes Problem. Die Entstehung der Information zur Entstehung des Lebens und der Höherentwicklung ist nicht durch Zufall möglich, sondern wird von dem Schöpfer geschaffen, der die Schöpfung plant, erschafft und lenkt. Dies erinnert an Augustinus, nach dem Gott die einen Schöpfungsobjekte vollständig, die anderen in einem „potenzialen latenten Zustand“, das Leben und die Organismen geschaffen hat, die auf günstige Bedingungen der Umwelt zur vollen Entfaltung warten, die fähig sind, sich weiter zu entwickeln gemäß des göttlichen Plans (siehe auch Kap. 1).37 Schöpfungswissenschaftler bezeichnen diese Vorstellung als „theistische Evolution“.
4. Abstammung und Sonderstellung des Menschen
Die heutige Hypothese der Abstammung des Menschen nimmt den Ägyptopithecus aus dem Oligozän vor 34 Mio. Jahren an. Die Stammformen der Menschenaffen und des Menschen, der Homoidea, werden an der Wende vom Oligozän zum Miozän vermutet. Innerhalb dieser Gruppe zeigen die Proconsulidae als eigene Gruppe eine hohe Variation. Eine Abstammung der Hominiden aus miozänen Menschenaffen ist aber fossil nicht belegt. Die frühen Hominiden-Fossilien nach der angenommenen Abspaltung vom Menschenaffen zum Menschen zeigen viele Arten und Gattungen. Man zählt 5 Gattungen in 15 Arten, von denen die Australomorphen eine ausgeprägte Radiation vom Pliozän zum Pleistozän zeigen. Ihr Gehirn ist menschenaffenähnlich, aber eine Werkzeugherstellung ist ungewiss. Die mosaikartige Verteilung von menschenaffenähnlichen und menschenähnlichen Merkmalen ermöglicht keinen Stammbaum; denn keine der heute bekannten Formen zeigt trotz der Fülle an Fossilien die Merkmale, die für eine Menschwerdung erforderlich sind.
Bei der Menschwerdung wurde jahrzehntelang ein baumlebender, hangelnder Menschenaffe angenommen, dessen Extremitäten keine Anpassung an zweibeiniges Gehen zeigen. Für eine Stammbaumkonstruktion wurden die vier Hauptmerkmale, aufrechter Gang, großes Gehirn, eine mit der Ernährung zusammenhängende Gesichtskonstruktion und die Befähigung zur Werkzeugherstellung als wesentlich erachtet. Es werden verschiedene Menschenaffen diskutiert, die in Frage kommen: Australopithecus africanus, A. habilis, A. garhi und A. afarensis vermutlich vor 3,1 Mio. Jahren.38 Sie zeigen für die Gehirngröße eine Variation von 280 bis 750 cm3 gegenüber den heutigen Menschen von 800 bis 2.000 cm3. Ende Miozän wird ein hypothetischer Übergang vom Menschenaffen zum Menschen vermutet (Affen-Mensch-Übergangsfeld). Die Gattung Homo tritt vom Plio- zum Miozän in drei Grundformen, Homo ergaster/erectus, H. neanderthalensis und H. sapiens, als echte Menschen auf. Der erste Mensch am Übergang vom Pliozän zum Altpliozän ist der Homo erectus. Diese Menschengruppe zeigt in Körpergestalt, Schädelmorphologie und Gehirngrößen spezielle Ausprägungen der Merkmale, die u. a. auf Klimaanpassungen zurückgeführt werden können. Wie die Australopithecinen im Pliozän erlebte die Gattung Homo eine starke Radiation. Der vor kurzem entdeckte zwergenartige Homo floresiensis zeigt wiederum, dass die Intelligenz nicht mit der Gehirngröße korreliert ist: 380 cm3, ca. 25% unseres Gehirns.39 Die Werkzeugherstellung ist zwar für den Menschen typisch, ist aber auch bei Tieren wie Schimpansen und Vögeln bekannt.
Von der Homo ergaster/erectus-Gruppe bildet H. ergaster eine frühe grazile Form im Nahen Osten, die robustere asiatische Form ist der H. erectus. Ihn kennzeichnet die Gehirngröße von 800 bis 1100 cm3, ein dem heutigen Menschen ähnlicher Knochenbau und ausgefeilte Jagdtechniken. Auch ist die Verwendung des Feuers nachgewiesen. Er soll vor 2 Mio. Jahren im Pliozän/Pleistozän in einer großen Ausbreitungswelle Afrika als H. ergaster und Asien als H. erectus besiedelt haben. Zu Mischformen, die vor 1 Mio. Jahren im Mittelpleistizän eine zweite Besiedlungswelle hervorgebracht haben, werden z. B. Homo antecessor und Homo heidelbergensis gerechnet. Der Neandertaler Homo neanderthalensis wurde wegen seiner großen Ähnlichkeit mit dem heutigen Menschen als Homo sapiens neanderthalensis bezeichnet, jedoch scheinen mitochondriale DNA-Untersuchungen für eine Abspaltung bereits vor 0,6 Mio. Jahren vom H. sapiens zu sprechen. Ihn kennzeichneten u. a. Totenbestattung, kulturelle und musische Leistungen, wie sich in einem Musikinstrument zeigt. Das große Gehirn wird auf eine Kälteanpassung im Zusammenhang mit der Eiszeit auf die Bergmannsche Regel zurückgeführt (größere Organismen mit relativ zur Masse kleinerer Oberfläche haben kleinere Wärmeverluste). Mit dem Homo sapiens, der wie der Neandertaler im Mittelpliozän erstmals nachgewiesen wurde, kam es zur dritten Besiedlungswelle mit der Ausbreitung in der Alten Welt vor 100.000 Jahren und in der Neuen Welt vor 30.000 Jahren. 0,4 Mio. Jahre alte Speere ähneln modernen Wurfspeeren. Herausragend ist die 200.000 Jahre alte Kunst, die dem Homo heidelbergensis zugeordnet wird, wie auch ein Mondkalender. Acheulienäxte mit hoher Ästhetik zeigte bereits Homo erectus. Schmuck und Flöten mit diatonaler Tonskala wie auch eine löwenköpfige Figur aus Mammutelfenbein werden dem Neandertaler zugeschrieben. Der Mensch zeigt sich auch als Künstler in den mächtigen Höhlenmalereien. Die Wissenschaftler hatten Schwierigkeiten, sie als menschlichen Ursprung anzuerkennen, da sie sich durch die Abstammungshypothese festgelegt hatten. Zeugnisse der Gottesverehrung in Tempeln, Altären und Totenkult zeugen von einem gläubigen und bewusst fühlenden Wesen. Gewaltige, rätselhafte Bauten (wie Pyramiden, „Unterwasserpyramiden“ vor der japanischen Küste, die Nazca-Kultur Südamerikas u. a.) aus der Urgeschichte der Menschheit, die sich in die momentane Menschheitsgeschichte kaum einordnen lassen, zeugen von den einzigartigen Fähigkeiten des Menschen. Die Archäologie steht vor vielen völlig ungeklärten Rätseln.40
Als Ursprungsland des Menschen gilt Afrika, aber auch der Nahe Osten und Eurasien werden diskutiert. Nach den Ergebnissen der mitochondrialen DNA-Untersuchung steht neben einem polyzentrischen Abstammungsmodell ein monozentrisches Modell „Arche Noah“ in Einklang mit der „Mutter Eva-Theorie“, nach der alle heute lebenden Menschen von der „Mutter Eva“ abstammen, die vor 200.000 Jahren gelebt haben soll.41
Aus dem hypothetischen Stammbaum des Menschen wurde ein undurchschaubarer Stammbusch. Daher kann man annehmen, dass ähnlich wie „Omne vivum ex vivo“ für alle Organismen gilt, der Mensch immer vom Menschen abstammt.
Der Paläontologe F. Schrenk, der als hervorragender Humananthropologe die Spuren der Vor- und Frühgeschichte erforscht, kommt zu dem Schluss: Die Frage nach dem Ursprung der Gattung Homo lässt sich mit den vorhandenen Fossilien allein auf anatomischer Grundlage nicht beantworten.42
Die Sonderstellung des Menschen in seinen einzigartigen Eigenschaften ist bei Naturwissenschaftlern unbestritten. Schon im Zeugungsakt „erkennen sich“ Mann und Frau von Angesicht zu Angesicht, in Liebe zum Mitschöpfertum berufen. Im ganzen Tierreich ist dergleichen nicht bekannt. Mit der Befruchtung der Eizelle durch die Samenzelle beginnt das Leben des Menschen.43 Berührt die Samenzelle die Eizelle, breitet sich über die Oberfläche der Eizelle ein Aktionspotenzial aus. Als entscheidender Moment lässt sich die Membranverschmelzung von Ei- und Samenzelle feststellen. Es folgt eine Ausschüttung des Inhalts zytoplasmatischer Vesikel in den perivitellinen Spalt (kortikale Reaktion). Schlagartig wird die Struktur der Zona pellucida verändert (Reaktion der Zona pellucida) mit Bildung des Polyspermieblocks. Nur eine einzige Samenzelle gelangt durch die erfolgreiche Membranfusion in die Zelle. Ab diesem Zeitpunkt wird die Diploidie durch die zwei Vorkerne der Keimzellen angebahnt, die schließlich, morphologisch ununterscheidbar, aufeinander zuwandern und das Geschlecht des neuen Menschen festlegen. Nur die Samenzelle kann das männlich bestimmende y-Chromosom oder das weiblich bestimmende x-Chromosom enthalten. In der dualistischen Sichtweise nach Hirnforscher und Nobelpreisträger Sir J. C. Eccles kann die Umsetzung vorhandener Erbinformation mit der Erschaffung eines geistigen Prinzips Seele verknüpft werden, die bei der einzigartigen leiblich-geistig-seelischen Entwicklung des Menschen mitwirken kann.44 Das Verschmelzungsprodukt von Ei- und Samenzelle, die Zygote, ist der Mensch im frühen Stadium. Schon die Eizelle des Menschen und die menschliche Zygote sind unverwechselbar mit Zygoten und Eizellen von Säugetieren. Die weitere Entwicklung des Embryos ist in jeder Phase als typisch menschlich beobachtbar, in der sich der Mensch „als Mensch entwickelt“ und nicht zum Menschen; denn „er ist Mensch in jeder Phase seiner Entwicklung von der Befruchtung an", wie der Embryologe Erich Blechschmidt in seiner Forschung erkannt hat. Ab dem 4. Tage verursacht der Mensch als Synzytiotrophoblast die Produktion des „human chorionic gonadotropin“ (HCG), das von der Mutter an den Urin abgegeben wird und den frühesten Schwangerschaftsnachweis nach 4 Tagen ermöglichen kann. Bereits in frühen Stadien mit der menschlichen Keimesentwicklung werden Kopf und Hände sehr auffällig; denn das eigentliche Werkzeug sind die „geistgelenkten Hände des Menschen“.45 Sie sind kein für den „Kampf ums Dasein“ ausgerüstetes Organ, wie man es hochspezialisiert bei Tieren findet, sondern ein für die Möglichkeiten des Werdens eines „Was-noch-nicht-ist-Hervorbringen-Könnens“, des kreativen Tuns.46
Die Ontogenese verläuft ohne Adultmerkmale vermeintlich tierischer Vorfahren (Ernst Haeckel), wie die Ergebnisse der Embryologie lehren.47 Alle sogenannten rudimentären Organe, wie „Schwimmhäute“, „Fell“ (Lanugo-Behaarung, auch bei Affen vorhanden) und „Dottersack“ treten nur vorübergehend auf und sind aus konstruktiven Gründen erforderlich. Die Ontogenese hat also keine tierischen Durchgangsstadien sondern ist typisch menschlich. Auch die sogenannten „primitiven Reflexe“ sind nach Blechschmidt ein Spiegelbild koordiniert ablaufender Differenzierungsvorgänge in der Form und in der Funktion. So sind die bogenförmigen Strukturen der frühen Embryonalentwicklung Beugefalten und keine Kiemenspalten einer Stammesgeschichte. „Die Entstehung eines Organs ist bereits der Beginn seiner Funktion“, fand Blechschmidt.48 Die Ohrmuskulatur z. B. entsteht beim nur 15 mm großen Embryo in Dehnungsfeldern. Die Anlagen für den aufrechten Gang und das große Gehirn des Menschen erschweren die Geburt, und der enge Geburtskanal erfordert komplizierte Drehungen des Kindes. Die herausragende Kopfgröße ist seit dem Auftreten des ersten Menschen nachweisbar. Die anatomischen Gegebenheiten empfehlen eine Geburtshilfe. Das Baby ist Nestflüchter und kommt mit offenen Augen und weit fortgeschrittener nervlicher und muskulärer Ausformung zur Welt. Es lernt im ersten Jahr die aufrechte Haltung, die durch Lebensäußerungen der Gruppe (der Familie) bestimmt wird.49 Diese „soziale Uteruszeit“ (Portmann) benötigt den internen Sozialkontakt von Mutter und Vater. Der Mensch erwirbt die Sprache als Instrument der „sozialen Vererbung“ in jahrelangem Lernen. Kein Säugetier zeigt eine 12-jährige Hemmung der geschlechtlichen Entwicklung wie der Mensch, sondern sie verläuft beim Säugetier ohne Verzögerung bis zur Geschlechtsreife. Dank des Meiosehemmfaktors verharren die Oozyten im Diktyotänstadium bis zur Pubertät. Die Spermatogenese erfolgt erst mit der Pubertät. In der Phase bis zur Pubertät müssen in Reflexion mit der Umwelt die vielen Anlagen der Sprache, des Verhaltens (auch der Scham) und der Talente entwickelt werden. Zur Entwicklung der Wortsprache in der sozialen Umgebung mit einer „vollen menschlichen Wertigkeit“50 sind die Zuwendungen der Mutter und auch des Vaters unentbehrlich. Eine künstliche Verkürzung der Ruhejahre der geschlechtlichen Entwicklung durch Frühsexualerziehung geht zu Lasten der geistig-seelischen Entwicklung. Es sind zahlreiche recht gut dokumentierte Extremfälle in den „Wolfskindern“ und im Kaspar Hauser-Phänomen mit den Folgen des Ausbleibens der Erziehung bekannt. Über den Ursprung der Würde des Menschen erkennt Robert Spaemann, dass der Mensch die Würde der Person von Anfang an besitzt, da er sie nicht erwerben muss. Die geistigen Kräfte wie Kreativität, Vernunft, Kunstfertigkeit, Gemüt, Gewissen machen den Menschen allen Lebewesen weit überlegen und begründen die materielle menschliche Würde.
5. Alternativen zur Evolutionslehre
Die wissenschaftlich bedenkliche grundsätzliche Einschränkung zahlreicher ungelöster Probleme ausschließlich mit atheistisch-materialistischen Forschungsmethoden lösen zu wollen, nötigt besonders in jüngerer Zeit Wissenschaftler Alternativen in Erwägung zu ziehen.51
5.1 Das Intelligent-Design-Konzept
Eine moderne Alternative ist das besonders in den USA verbreitete und von bekannten Wissenschaftlern wie Behe52 unterstützte „Intelligent Design“-Konzept (ID).53 Einstein sagte einmal: „Im unbegreiflichen Weltall begegnet uns eine unendlich überlegene Vernunft“, d. h. auch, an den Strukturen, Gesetzen und ablaufenden Prozessen in den Organismen lässt sich ein überlegener Vernunft-begabter intelligenter „Designer“ als Urheber erkennen. Philipp Johnson, der 1993 „Darwin on Trial“ verfasst hatte, schrieb 1998 „Intelligente Ursachen können bewirken, was nicht intelligente Ursachen nicht können“.54 Hinweise lassen einen zugrunde liegenden Plan erkennen in für uns als Zufälligkeiten oder erprobten Gesetzen erscheinenden Phänomenen. Michael Ruse stellte fest: „Wir untersuchen Organismen – mindestens ihre Teile – als wären sie erschaffen, als wären sie entworfen („designed“) worden und dann versuchen wir ihre Funktion herauszufinden.“55 Die Hinweise auf eine solche Planung heißen „Design-Signale“. Solche können wir in den verschiedenen Komplexitäten erkennen:
5.1.1 „Irreduzible Komplexität“
In der „irreduziblen Komplexität“ genügt es, einen beliebigen Teil des Ganzen wegzulassen oder zu inaktivieren und die Funktion ist vollständig zerstört. Es wird daher angenommen, dass solche Teile in einer einzigen Generation entstanden sind, da die Zwischenstadien eines stufenweisen Aufbaus wegselektiert worden wären. Hierzu lässt sich der Bakterienmotor (siehe Kap. 3) aufzählen.56 Ein einziges defektes Gen des aus mindestens 40 Genen aufgebauten Motors genügt, um die Funktion auszuschalten. Viele Beispiele zeigen die Analogie der menschlichen Ingenieurskunst in den Konstruktionen lebender Organismen und werden umgekehrt heute für wirtschaftliche Zwecke nachgeahmt. Werner Gitt schreibt z. B. über die Vögel: „In der Tat ist die navigatorische Leistung, … die ohne Kompass und Karte und bei sich ständig ändernden Bedingungen von Sonnenstand, Windrichtung, Wolkendecke und Tag/Nacht-Rhythmus geschieht, ein Wunder ohnegleichen“.57 Der Goldregenpfeifer z. B. fliegt 4500 km im Non-stop-Flug von Alaska zu den Hawai-Inseln. Dem Vogel stehen als Treibstoff 70 g Fett zur Verfügung. Im Einzelflug wäre nach 72 Stunden das gesamte Fettpolster verbraucht und 800 km vor dem Ziel würde der Vogel ins Meer stürzen. Beim Flug in Keilformation spart er aber 23% Energie und erreicht sein Ziel. Eine minimale Abweichung vom Kurs würde bedeuten, dass der Vogel auf dem offenen Meer rettungslos zugrunde gehen müsste. Die exakte Einhaltung der Zugrichtung ist also ohne Einschränkung erforderlich und konnte nicht stufenweise erprobt werden, sondern es muss der Flugplan mit seinen Bedingungen in den Erbanlagen programmiert worden sein, den der Vogel unbewusst abarbeitet. Von den zahlreichen Komponenten, die hier zusammenwirken, darf keine ausfallen.
5.1.2 „Spezielle Komplexität“ (Luxurieren)
Viele Lebewesen zeigen Eigenschaften, die eine spielerische Komplexität aufweisen, ein „Luxurieren“, das nicht notwendig wäre, um einen bestimmten Zweck zu erreichen. Sieht man sich die kultivierte Balz der Laubenvögel an, so fragt man sich, ob eine einfachere Konstruktion nicht energiesparender, ökonomischer und zweckmäßiger wäre, als so die knappe Energie zu vergeuden: Der Rotlaubengärtner (Amblyformis subalaris) aus den Bergen Südost-Neuguineas baut eine Laube, die nach Goldwin das schönste Bauwerk darstellt, das je von einem Vogel geschaffen wurde. Es wurde bei seiner Entdeckung für ein vom Menschen gebautes Kinderspielzeug gehalten. Um ein „Baum“stämmchen werden dürre Ästchen 50 cm hoch horizontal angelehnt. Die Lücken zwischen den Zweigen werden mit Moos ausgefüllt, wodurch eine Säule entsteht. Von den Seiten werden Ästchen und Zweiglein so gelehnt, dass ein schräges Dach entsteht. Diese Hütte hat zwei Eingänge. Der Vorplatz wird mit gelben, roten, blauen Beeren und gelben Blumen belegt. Farbige Blüten, Käferflügel vervollständigen den schmucken Bau.58
Auch die komplizierte Verständigung der Honigbiene Apis mellifica darf man hier erwähnen. Ihre Richtungs- und Entfernungstänze folgen mathematischen Funktionen. Sie finden sich bei anderen Bienen nicht oder nur rudimentär.
5.1.3 „Potenzielle Komplexitäten“
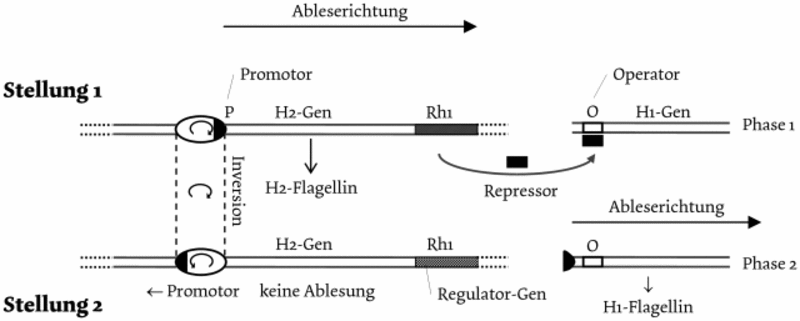
Als Ergebnis einer Selektion lassen sich „potenzielle Komplexitäten“ kaum ansehen. Hier liegen Programme vor, die nur im Bedarfsfalle eingesetzt werden, also in Reserve liegen, z. B. als Überlebensstrategie bei Salmonella typhi.59 Hier können H-Gene zwei verschiedenartige Strukturen hervorbringen. Die Primärstruktur des Flagellins, an dem die Antikörper angreifen können, wird durch zwei Gene bestimmt, von denen immer nur eins von der messenger RNA zu Proteinsynthese abgelesen werden kann (Abb. 4). Durch eine spontane Inversion der Ablesungsrichtung der DNA-Sequenz 10-4 pro Zellteilung wird dann z. B. nicht mehr das H2-Gen abgelesen, sondern das H1-Gen, das jetzt nicht mehr durch ein Repressorprotein am Operator am Ablesen des H1-Gens gehindert wird. Wenn sich die Antikörper auf die Geißelstruktur des H2-Flagellins der Salmonellen eingestellt hatten und den Bakterien die Vernichtung droht, kann durch Umschaltung der Ableserichtung das H1-Flagellin gebildet werden, gegen das keine spezifischen Antikörper vorliegen. Das Immunsystem braucht für die Einstellung auf H1-Flagellin Zeit, in der sich die Bakterien zu einer Gefahr für den Wirt vermehren können. Solche vorsorgenden in Reserve befindlichen Schutzeinrichtungen finden sich auch im Genetischen Code. Durch die sogen. Degeneration ist er ferner so organisiert, dass bei einer Mutation das DNA-Triplett durch ein anderes DNA-Triplett ersetzt werden kann, das die gleiche Aminosäure codiert. Der Genetische Code ferner ist so organisiert, dass bei einer Mutation zu einem anderen Triplett eine andere Aminosäure im Protein auftritt, die aber chemisch ähnlich ist, z. B. auch eine basische, saure oder neutrale (aliphatische oder aromatische) Molekülgruppe trägt. Hierdurch wird der Schaden z. B. einer möglichen Inaktivierung eines Enzymproteins vorsorglich begrenzt. Eine potenzielle Komplexität besitzen auch die Reparatur- und Regulationssysteme.
5.2 Die Schöpfungswissenschaft als Alternative zur Evolutionstheorie
Hier wird die Frage gestellt, wie sich die bereits vorgefundenen Naturerkenntnisse in einem biblischen Rahmen deuten lassen, wenn die Bibelberichte als inspirierte Wahrheit zugrundegelegt werden.60 Umgekehrt lassen sich Forschungsprogramme auf der Basis der Bibelberichte ausarbeiten. Das Schöpfungsmodell der Schöpfungswissenschaft steht dem Evolutionsmodell der Evolutionstheorie gegenüber. Es gilt auch, dem reinen Materialismus durch eine bibelorientierte Forschung entgegenzutreten, um Engführungen in der Wissenschaft zu vermeiden. Naturforscher bestätigen, dass Bibelberichte durchaus anregend sein können. Von Fernstehenden wird häufig eingewendet, dass Erklärungen für Ursachen kurzschlüssig unmittelbar auf den Schöpfer zurückgeführt werden und damit eine Ursachenforschung lahmlegen. Dem widerspricht sowohl die Genesis61 als auch das Tridentiner Konzil, das ausdrücklich aufforderte, die Schöpfung zu erforschen. Forschung bedeutet aber immer auch ein Infragestellen von als gesichert geltenden Tatsachen.
Die Schöpfungswissenschaft geht von einem geschaffenen Plan aus, der zielgerichtet zu „Passformen“ führt, entsprechend den Bibelzitaten: „Noch ehe ich dich im Mutterleib formte, habe ich dich ausersehen…“62 spricht der Herr zu seinen Propheten. „Als ich geformt wurde im Dunkeln, kunstvoll gewirkt in den Tiefen der Erde, waren meine Glieder dir nicht verborgen… Schon meine unfertigen Teile sahen deine Augen und in deinem Buch waren sie alle eingetragen“.63 Alles, was er geschaffen hat, schützt und lenkt Gott durch seine Vorsehung „sich kraftvoll von einem Ende bis zum anderen erstreckend und alles mild ordnend“.64 Die Realisierung, die Ausformung biologischer Objekte, erweckt den Eindruck einer Selbstorganisation („self assembly“), wie man bei der Bildung von Ribosomen, den Organellen der Proteinbiosynthese, und der Bakteriophagen (Bakterienviren) aus ihren Bestandteilen im Reagenzglas beobachten kann. Augenblickliche Realisierung ist möglich; denn der Herr sprach und sogleich geschah es. „Er gebot und alles war da“65 oder plangemäße Werdeprozesse wie in der Keimesentwicklung. Triebfeder der Schöpfungsakte, „creatio ex nihilo“ ist das „Fiat“ – „Es werde...“.66 Aufgabe der Schöpfungswissenschaft ist es, mit wissenschaftlichen Methoden den Plan der Schöpfung zu ergründen und seine Realisierung aufzudecken. Wie erwähnt, hat auch Augustinus Vorstellungen entwickelt von einem „potenzialen, latenten Zustand“ der Schöpfung, der nur auf günstige Bedingungen der Umwelt zur vollen Entfaltung warte.67
Die Schöpfungswissenschaft arbeitet mit allgemein anerkannten Methoden, jedoch in einem biblischen Deutungsrahmen, d. h., dass der Aktualismus (oder Uniformitarismus), der Grundlage der Lehre Darwins ist, abgelehnt wird. Wie berechtigt dies ist, zeigen geologische Arbeiten, in denen die Prinzipien des Aktualismus nicht angewendet werden können, z. B. bei der Bildung von Bändereisenerz. So schreibt Altermann: „Für die BIFs fehlt es bis heute an einheitlich gültigen Sedimentationsmodellen. Eine aktualistische Herangehensweise ist offensichtlich nicht anwendbar, da solche Sedimente seit dem Proterozoikum nicht mehr gebildet wurden“,68 oder Klemm schreibt: „Die außerordentliche Konzentration von Eisen kann durch ein uniformitaristisches Fällungsmodell schwerlich erklärt werden“.
Der Begriff des „Grundtyps“, der auf Frank L. Marsh zurückgeht, ist ein wichtiges Forschungsobjekt. Er beruht auf dem Konzept der Gruppen von Varianten einer Art oder Gattung, evtl. einer Familie, die auf die Bibelstellen Bezug nehmen: „Gott bildete … jeweils nach ihren Arten“.69 Alle Individuen, die direkt oder indirekt durch Kreuzungen miteinander verbunden sind oder nur durch den Beginn einer Embryonalentwicklung charakterisiert sind, gehören zu einem Grundtyp.70 Bei Mikroorganismen, wie Bakterien, sind es ähnliche Genfolgen auf dem Bakterienchromosom (siehe Kap. 2). Innerhalb eines Grundtyps sind Übergänge möglich. Übergänge zwischen den einzelnen Grundtypen, d. h. untereinander, sind aber nicht bekannt. Die Variabilität der Individuen innerhalb eines Grundtyps ist experimentell nachweisbar und deckt sich mit dem Begriff „Mikroevolution“ (Kap. 2).71
Die Grundtyptheorie wird gestützt durch Experimente, die von der Schweizer Firma CIBA in den 1980er Jahren durchgeführt wurden: In diesen wurden in einem elektrostatischen Feld Eier der Regenbogenforelle, Samen von Mais und Sporen vom Wurmfarn ausgesetzt. Dabei entstanden Abweichungen der heutigen Merkmalsausprägungen, die den Ahnenformen entsprachen. Die Forellen ähnelten in Gestalt und Verhalten Wildformen, der Mais zeigte büschelförmigen Kolbenwuchs und die Farnpflanzen erinnerten an 300 Mio. Jahre alte Farnfossilien. Diese Ergebnisse können auf Interaktionen der elektrostatischen Felder mit morphogenetischen Feldern erklärt werden. Erbanlagen früher Formen sind also noch präsent und können abgerufen werden, z. B. die Aktivierung ruhender Gene und Inaktivierung aktiver können die frühen Formen verständlich machen.72
Ein weiteres Tätigkeitsfeld der Schöpfungswissenschaft bilden Fossilgeschichte und Sintflut73, deren chronologischer Ablauf als Folge eines Kometeneinschlags dargelegt wird.74 Für diese globale Katastrophe sind Bausteine einer biblisch-urgeschichtlichen Geologie („Flutgeologie“) in einem Kurzzeitraum zu erarbeiten. Die Fossilien sind das Ergebnis eines gewaltsamen Todes und müssen daher zeitlich nach dem Sündenfall eingeordnet werden. Die Gesteinsbildung muss rasch erfolgt sein. Die Sedimentgesteine können weltweit nach dem Überlappungsprinzip in einer Fossilgeschichte eingeordnet werden,75 wie dies in der Wissenschaft bereits vor dem Durchbruch der Evolutionstheorie 1750 bis 1850 noch geschah. So haben die Formationen der geologischen Zeittafeln nichts mit der damit verbundenen Zeitdatierung zu tun. Für Kurzzeiträume gibt es heute konkrete Anhaltspunkte. Der 1980 ausgebrochene Vulkan Mt. St. Helens zeigte schnelle Ablagerungen der Schlammströme mit bis zu 30 m hohen Felswänden, wie der Little Grand Canyon innerhalb von Stunden.76 Für einen kurzen Zeitraum in der Vergangenheit treten Norman und Setterfield in den USA seit 1987 ein, die mit einer „dynamischen Zeitbestimmung“ aufgrund einer zeitabhängigen Verringerung der Lichtgeschwindigkeit nach mathematischer Auswertung aller Bestimmungen der Lichtgeschwindigkeit seit Olaf Römer Aufsehen erregt haben.77 Eine Inkonstanz der Lichtgeschwindigkeit wird von Physikern diskutiert, wie der Physiker João Magueijo dargelegt hat.78 Nach Studien der gesamten Menschheitsgeschichte kam Brandt zu dem Schluss, dass diese nicht länger als 10 000 Jahre gewährt haben kann.79
Bereits im 17. und 18. Jahrhundert haben die Forscher Burnet (1684) und Liliental (1756) die Aufspaltung der Landmasse infolge der Sintflut zu den heutigen Kontinenten biblisch begründet: „Einer hieß Peleg, weil zu seiner Zeit die Erde zerteilt wurde“.80 Diese wurde im 20. Jahrhundert von Alfred Wegener in der Kontinentalverschiebungstheorie nachgewiesen, fand aber zunächst bei aktualistisch eingestellten Wissenschaftlern Ablehnung. Heute ist sie allgemein anerkannt.
Die Sintflutforschung hat in der Feststellung, dass die biblischen Daten zum Bau der Arche so gestaltet sind, dass sie optimaler Schwimmstabilität bei gleichzeitig minimalem Material- und Arbeitseinsatz entsprechen, einen großen Erfolg errungen.81 Unter den diskutierten Sintflutmodellen im Kurzzeitraum ragt das Kambrium-Perm-Modell Joachim Schevens heraus.82 Der mit der Flut schlagartig einsetzende Fossilreichtum ist im Kambrium nachweisbar. Die rätselhaften Worte bei Ezechiel „Mit den Bäumen von Eden unter die Erde hinab“83 bezieht Scheven auf die Vegetation der vorsintflutlichen Erde. Danach sind diese Bäume, Bärlappbäume (Sigillaria), Erzeuger der Steinkohle mit typischer Rinde, die mit Zentralzylindern durch Leitbündel verbunden ist, wobei der übrige Raum mit Luft gefüllt ist (Fossil Grove in Glasgow 1983). Sie bilden riesige Schwimmwälder, die in rasch absinkende Ablagerungströge eingeschwemmt wurden und in der Tiefe gleich unter hohen Druck gelangten. Diese belegbare Theorie der Steinkohlenentstehung liefert einen wesentlichen Beitrag zur Schöpfungslehre.84 Die Schulgeologie hat nichts Überzeugenderes zur Steinkohlenbildung zu bieten. Scheven entwickelte auch eine interessante Theorie der Braunkohlenentstehung im Tertiär.85
Hinweise auf den Untergang der Dinosaurier, die häufig im Wasser leben, gibt eine Stelle aus den Psalmen: „Du trennst das Meer durch deine Kraft und zerbrichst die Köpfe der Drachen im Meer“.86 Als Ergebnis des Trockenfallens der nachsintflutlichen Flachmeere infolge des Abfließens des Wassers von den Kontinenten verlieren die Dinosaurier ihren Lebensraum und gehen schnell zugrunde, wie ihre Skelette, Eigelege und Fußspuren von einer augenblicklichen Zudeckung mit Sand und Schlamm bezeugen. Diese ökologische Theorie Schevens ist nicht nur sehr einsichtig und wurde durch neue Funde bestätigt (in Münchehagen-Rehberg 1982 bekannt geworden), sondern entspricht dem aktuellen Forschungsstand (Kap. 1).
Die wenigen angeführten Beispiele der Alternativen zur Evolutionstheorie lassen erkennen, dass diese Aussicht haben, in Konkurrenz zum Evolutionsmodell treten zu können. Die wissenschaftliche Redlichkeit verlangt, dass eine Fülle von Befunden ohne Erklärung nach der Evolutionstheorie nicht verschwiegen, sondern in den einschlägigen Fachbüchern für Denkanstöße dargelegt werden, z. B. ein Fährtenzug von menschlichem Aussehen im Karbon, der als „unecht“ oder als „Amphibienfährte“ abgetan wird oder die beschlagenen Feuersteine von Aurillac bzw. Lüttich, von Breuil (1910) und Überbaier (1924) im Paläozän des Tertiärs vor ca. 58 Mio. Jahren, die als „Zufallsprodukte der Natur“ erklärt werden.87
6. Über die gesellschaftliche Bedeutung der Evolutionstheorie
Die Lehre Darwins hatte tiefgehende Auswirkungen auf die verschiedensten Bereiche der Gesellschaft. Die Zufallslehre führte zu einer Abkehr von Gott. Der „Kampf ums Dasein“ hatte nicht nur die Umgangsformen rauer gestaltet, sondern den wirtschaftlichen Kapitalismus bis zum „Wolfskapitalismus“88 verhärtet. „Wo Leistung verlangt wird, hat die Bergpredigt nichts zu suchen“, meinte ein Unternehmer bei einer Veranstaltung der Führungskräfte einer Diözese. Vor der Gefahr einer atheistischen Gesellschaft hatte schon Augustinus gewarnt: Eine Gesellschaft ohne Gott ist bestenfalls eine gut organisierte Räuberbande.89 Die Verwilderung der Sitten auf allen Gebieten in den Massendemokratien der westlichen Länder wird schon gar nicht mehr wahrgenommen, wie sich zeigt, wenn gläubige Moslems Integrationsversuchen Widerstand leisten.
6.1 Marxismus und Evolutionstheorie
In Deutschland sah der berühmte Arzt Rudolf Virchow 1877 in München in der Evolutionstheorie „eine ungemein bedenkliche Seite und dass der Sozialismus mit ihr Fühlung aufgenommen hat, wird ihnen hoffentlich nicht entgangen sein“.90 Anders Ernst Haeckel, der für Darwin energisch eintrat und zwei einander schroff gegenüberstehende Parteien sah: „Auf der einen Fahne der progressiven Darwinisten stehen die Werte „Entwicklung und Fortschritt“, und aus dem Lager der konservativen Gegner Darwins tönt der Ruf „Schöpfung und Spezies“.91 Marx und Engels begrüßten die Lehre Darwins besonders angesichts der himmelschreienden Zustände im aufkommenden Kapitalismus, der vom Recht des Stärkeren bedenkenlos Gebrauch machte. Der Russe Timirjasew erläuterte im zaristischen Russland die Entwicklungslehre in ihrer Bedeutung für den Sozialismus in der Schrift „Charles Darwin und Karl Marx“.92 Die Tab. 1 stellt die Lehren von Darwin und Marx gegenüber mit verblüffender Übereinstimmung. Den Organismuspopulationen Darwins mit ihrer Veränderlichkeit trotz einer weitgehenden Konstanz in der Theorie entsprechen die Produktionsverhältnisse in einer Gesellschaftsordnung mit einer stabilen Klasse. „Sie bilden die ökonomische Struktur der Gesellschaft, die reale Basis, worauf sich ein juristischer und politischer Überbau erhebt“ (Marx und Engels), der sog. Ideologische Überbau. Aber angesichts der sich häufenden gesellschaftlichen Missstände entstehen Ideen zu ihrer Veränderung. Dieses dynamische Prinzip zur Veränderung bilden die Umwelt nach Darwin und die Produktivkräfte nach Marx, die von den arbeitenden Menschen mit den Produktionsmitteln, der Technik und der Bodenschätze etc. gebildet werden. Die Entstehung des Neuen erfolgt einerseits durch Umweltveränderungen, die zu neuen „Spielarten“ führen, weil die bestehende Population mit den neuen Umweltverhältnissen in „Widersprüche“ (Marx) gerät. Andererseits entstehen neue Produktionsverhältnisse, wenn sich die Produktivkräfte verändern, wodurch diese mit den veränderten Produktivkräften in Gegensatz, „Widersprüche“ nach Marx, geraten. Das Selektionsprinzip wirkt sich „richtunggebend“ aus, indem die vorherrschenden Organismen „im Kampf ums Dasein“ durch die neuen Spielarten verdrängt werden, die besser angepasst sind (Darwin), während nach Marx die alten Produktionsverhältnisse in einer Revolution durch neue Produktionsverhältnisse ersetzt werden, weil sie den neuen Produktivkräften, d. h. der neuen Klasse, besser entsprechen. „Auf einer gewissen Stufe“, lehrt Marx, „geraten die materiellen Produktivkräfte der Gesellschaft in Widerspruch mit den vorhandenen Produktionsmitteln, innerhalb der sie sich bisher bewegt hatten. Aus Entwicklungsformen der Produktivkräfte schlagen diese Verhältnisse in Fesseln derselben um. Es tritt eine Epoche der sozialen Revolution ein.“93 Ähnlich wie die neu angepasste „höher entwickelte“ Organismenpopulation an die neuen Umweltverhältnisse angepasst ist und sich etabliert, bilden die neuen Produktionsverhältnisse der neuen „Klasse" die neue „höher entwickelte Gesellschaftsordnung“. Dem Darwinismus entspricht also im Wesentlichen die später als „wissenschaftlicher Sozialismus“ bezeichnete Klassenkampftheorie von Karl Marx.
| Die auf Darwins Lehre beruhende Evolutionstheorie | Die auf Marx und Engels beruhende Theorie der Klassenkämpfe des wissenschaftlichen Sozialismus | |
| Altes Gleichgewicht | Organismenpopulation zeigt Artkonstanz mit Entstehung von Varietäten | Produktionsverhältnisse zeigen Klassenkonstanz mit Entstehung neuer Ideen, wirtschaftlicher, geistiger, gesellschaftlicher und juristischer Natur |
| Dynamisches Prinzip | Umwelt (Klima, Temperatur, Feuchtigkeit, Katastrophen usw.) | Produktivkräfte (ausgebildete, arbeitende Menschen und Produktionsmittel, Technik, Bodenschätze usw.) |
| Die Entstehung des Neuen | Neue Spielarten („sports“) entstehen, wenn sich die Umwelt ändert | Neue Produktionsverhältnisse entstehen, wenn sich die Produktivkräfte verändern |
| Die Organismen geraten mit der veränderten Umwelt in Gegensatz | Die Produktionsverhältnisse geraten mit den veränderten Produktivkräften in Gegensatz (= „Widerspruch“ bei Marx) | |
| Wirkung des Selektionsprinzips | Neue Spielarten verdrängen vorherrschende Organismen (auch durch Kampf ums Dasein) | Neue Produktionsverhältnisse verdrängen die vorherrschenden Produktionsverhältnisse (auch durch Revolution) |
| Neues Gleichgewicht auf höherer Ebene | Höherentwickelte Organismen herrschen; z. B. Folge: Fische, Lurche, Reptilien (und Vögel?), Säugetiere | Höherentwickelte Produktionsverhältnisse bestimmen die neue Klasse in den jeweiligen Gesellschaftsordnungen; z. B. Folge: (Urgemeinschaft) > Sklaverei > Feudalismus > Kapitalismus > Sozialismus (> Kommunismus) |
Tab. 1: Vergleich der Evolutionstheorie mit der Theorie des Klassenkampfes des wissenschaftlichen Sozialismus
Doch auch der Sozialismus, den man in der Sozialdemokratie als eine Fortentwicklung und Abschwächung des Marxismus sehen kann, zeigt in seinem Bestreben, allen Menschen von Anfang an gleiche Entwicklungsmöglichkeiten zu geben, damit auf dieser Grundlage der „Chancengleichheit“ die natürliche Auslese, der Konkurrenzkampf, entbrennen kann, Züge des Darwinismus. Aber da es nicht möglich ist, die Menschen gleich zu machen, kann der sozial Schwache nicht mithalten, wenn er die Mittel im Konkurrenzkampf zu obsiegen nicht besitzt, wie Grün meint.94
Zu bedenken ist, dass der Marxismus eine Reaktion auf unsoziales Verhalten von Unternehmern darstellt, das Marx vor allem in England studierte. Dabei entstand das Kommunistische Manifest und die Theorie von der „Ausbeutung als Aneignung des Mehrwertes durch den Besitzer des konstanten Kapitals“, d. h. vor allem der Unternehmen. Die Technisierung, z. B. des Webstuhls, hat in der Industriellen Revolution hier eine wesentliche Rolle gespielt. Der Verelendung der Arbeiterschaft hat sich Papst Leo XIII. 1891 in der Sozialenzyklika „Rerum novarum“ angenommen, die von Gewerkschaften und Arbeiterbewegungen aufgegriffen wurde. Auch heute zeigt sich in einer fortgeschrittenen Technik statt der Marktwirtschaft Ludwig Erhards eine kapitalistische Hochform des Kapitalismus, ein „Raubtierkapitalismus“ nach Peter Scholl-Latour. Aktionäre und Direktoren billigen sich zweistellige Gehaltserhöhungen zu bei gleichzeitiger Massenentlassung der Arbeitnehmer. Hier wird gleichfalls ein Sozialdarwinismus durch „survival of the fittest“ demonstriert.
6.2 Der Darwinismus und die Lehre des Christentums
Es hat nicht an Versuchen gefehlt, die Evolutionslehre in das christliche Lehrgebäude zu integrieren, wie auch an Versuchen nicht gespart wurde, Marxismus und Christentum, den Geist der Bergpredigt, miteinander zu harmonisieren. So hat Herbert Haag seine Schrift „Abschied vom Teufel“ damit begründet, dass die Evolutionstheorie die biblische Lehre vom Teufel nicht mehr zuließe. Karl Rahner lehrte, dass die Erfahrungstatsache des Werdens „Grundaxiom der Theologie“ sei. An die Stelle der Erlösung vom Tod tritt der „Tod als erlösendes Geschehen, als selektives Prinzip der Höherentwicklung“ (zit. n. Alma von Stockhausen). Auch von Seiten gläubiger Naturwissenschaftler werden Thesen verfochten, wie „Gott schafft durch Evolution“.95 Es stellt sich die Frage, ob der Standpunkt des Kompromisses mit der Makroevolution in der sog. „theistischen Evolution“ nicht „das Wort Gottes als absoluten Maßstab auflöst und dann zu einem theologischen Liberalismus, zur Theologie der leeren Kirchen, führt“.96 Die Auswirkungen der Evolutionslehre auf die christliche Lehre wurde u. a. von Reinhard Junker untersucht.97 Die wesentlichen Punkte seien hier aufgeführt:
1. Die Sünde, vor allem die Erbsünde, die die physischen und moralischen Übel erklärt (Pascal), ist nicht mehr denkbar; denn „Sünde“ ist nur ein Nebeneffekt eines auf Versuch und Irrtum basierenden Evolutionsprozesses. Damit entfällt die biblische Deutung des Übels und die Lösung des Theodizee-Problems.
2. Das Böse ist keine gottwidrige Macht, sondern dem Entwicklungsprozess eigen. Das „Gleichgewicht des Schreckens, des Fressens und des Gefressenwerdens“ ist der Motor der Entwicklung. So wird die Sünde ein Begleitphänomen der „Höherentwicklung“. Damit entfällt die biblische Ethik von Schuld und Sühne.
3. Die Erlösung ist keine Befreiung von der Sünde und Aussöhnung mit Gott durch den Sühnetod des eingeborenen Sohnes Jesus Christus. Die Zukunft kann nach Teilhard de Chardin „in einer Vollendung der Evolution mit dem Erreichen des Punktes Omega“ bestehen. Das Ertragen von Leid in der Kreuzesnachfolge Christi ist sinnlos. So erklären sich die Bestrebungen, Schwierigkeiten zu bewältigen, z. B. das Kind im Mutterschoß töten zu lassen oder in der Euthanasie einen Ausweg zur Lösung des Problems einer überalterten Gesellschaft zu sehen.
4. Der Mensch ist auf dem Weg vom Tier zum Menschen; denn er habe das Tier-Mensch-Übergangsfeld noch nicht vollständig verwirklicht. Damit bildet Jesus nur eine Evolutionsstufe. Für die Sonderstellung des Menschen ist kein Platz, wie sie heute gewöhnlich mit allem Nachdruck bestritten wird.98 So erklärt sich auch der Umgang mit Entwicklungsphasen des Menschen, obwohl der Mensch nach Erkenntnissen von Blechschmidt „in jeder Phase ganz Mensch“ ist.99 Die besonders von der Katholischen Kirche geforderte Disziplin in der Sittenlehre kann nach der Autonomen Moral nach Alfons Auer100 getrost der Entscheidung des Einzelnen überlassen werden ohne Ausrichtung des Gewissens auf die Lehre der Kirche, z. B. in der Frage der Empfängnisverhütung in der Königsteiner Erklärung.
5. Die unsichtbare Schöpfung von Engeln und Dämonen ist eine überholte Lehre aus dem „finsteren Mittelalter“ (Französische Aufklärung), als die großen Errungenschaften der Naturwissenschaften und der Technik noch unbekannt waren. Eine biblisch fundierte Lehre von Läuterung (Fegefeuer) oder Gottesferne (Hölle) ist veraltet und kann dem modernen Menschen, geschweige denn den sensiblen Kindern nicht zugemutet werden.
6. Der Tod ist nicht der Sold der Sünde, sondern Voraussetzung der Evolution mit den sich ablösenden Organismenpopulationen zur Höherentwicklung. Die Auferstehung erscheint als Illusion; denn Leid und Tod sind „Schöpfungsmittel“ der Evolution. Unvollkommenheiten sind der Evolution eigen und aus dem Zufallsmechanismus, aus dem „Versuch und Irrtum“ verständlich. Der Begriff von Gott als dem Inbegriff des Guten ist nur noch subjektiv, nur noch in unserem Herzen denkbar.101
7. Allein in der Materie und in den in ihr innewohnenden Gesetzmäßigkeiten gründet der Evolutionsprozess, ohne Einwirken eines Gottes in den Kosmos. Die immaterielle Welt, wie Mystik in kirchlich anerkannten Marien- und Engelerscheinungen von Fatima und Heilungswundern in Lourdes sind Sinnestäuschungen, psychosomatische Effekte oder Selbstheilungsprozesse. Für evolutionistisch eingestellte Theologen sind Wunder Unfug (z. B. Heroldsbach und Marienfried). Das gilt auch für die Wunder Jesu, die nach manchen Theologen als „legendär“ oder „legendarisch“ im Dienste der Verkündigung (Historische Bibelkritik) zu betrachten sind.
8. Die christliche Heilsaussage lässt sich nur im Lichte einer kosmischen Evolution sehen, die einen göttlich durchwirkten Prozess darstellt.
Die logischen Schlussfolgerungen aus der Evolutionslehre sind somit nicht für Christen akzeptabel. Es gibt keinen Kompromiss. Die Haltung des Christen steht in Übereinstimmung mit den naturwissenschaftlichen Ergebnissen, wenn die Mikroevolution als Tatsache und die Makroevolution als Arbeitshypothese angenommen wird in der Gewissheit, dass Wissenschaft, d. h. Vernunft einerseits, und Glaube, d. h. Offenbarung andererseits, einander nicht widersprechen, weil beide aus der Hand des Schöpfers stammen. Eine atheistische Öffentlichkeit aber reduziert die christliche Glaubenslehre auf die Besserung des Menschen, insbesondere in sozialen Fragen, und belässt Kirchen als Hilfskorps bei der Lösung sozialer Fragen, wie der Krankenpflege, Altenpflege und zur Stabilisierung der Gesellschaft, aber auch als Denkmalsschützer und Interpreten christlicher Kunst. Diese Betätigungsfelder werden von atheistischen Diktaturen wie Massendemokratien gleichermaßen geduldet.
6.3 Darwinismus als Grundlage der Rassentheorie des Nationalsozialismus
Es kann kein Zweifel bestehen, dass vom Nationalsozialismus die Thesen von „struggle for life“ und „survival of the fittest“ auf den Menschen für die Rassentheorie missbraucht wurden, wie z. B. eine Hitlerrede am 22. Juni 1944 an Offiziersanwärter belegt: „Die Natur lehrt uns bei jedem Blick in ihr Walten, dass das Prinzip der Auslese sie beherrscht, dass der Stärkere Sieger bleibt und der Schwächere unterliegt. Sie lehrt uns, dass das, was den Menschen dabei als Grausamkeit erscheint, weil er selbst betroffen ist oder weil er durch seine Erziehung sich von den Gesetzen der Natur abgewandt hat, im Grunde doch notwendig ist, um eine Höherentwicklung der Lebewesen herbeizuführen. Die Natur kennt vor allem nicht den Begriff der Humanität, der besagt, dass der Schwächere unter allen Umständen zu fördern und zu erhalten sei, selbst auf Kosten der Existenz des Stärkeren… Die Natur kennt in der Schwäche keinen Milderungsgrund,… im Gegenteil, die Schwäche ist der Grund der Verurteilung… Der Krieg ist also das unabänderliche Gesetz des ganzen Lebens, die Voraussetzung für die natürliche Auslese des Stärkeren und zugleich der Vorgang der Beseitigung des Schwächeren. Das, was dem Menschen dabei als grausam erscheint, ist vom Standpunkt der Natur aus selbstverständlich und weise. Ein Volk, das sich nicht zu behaupten vermag, muss gehen und ein anderes an seine Stelle treten. Ein Wesen auf dieser Erde wie der Mensch kann sich nicht dem Gesetz entziehen, das für alle anderen Wesen auch gültig ist… Seit es Wesen auf dieser Erde gibt, ist der Kampf das Unvermeidliche“.102 Damit ist nach Johannes Grün dem Urteil Erich Fromms zuzustimmen: „Der soziale und moralische Darwinismus ist ein romantisches Heidentum und gehört zu den gefährlichsten Elementen im Denken des letzen Jahrhunderts…Insofern Hitler überhaupt an etwas glaubte, so waren es die Gesetze der Evolution, die sein Handeln und speziell jede Grausamkeit rechtfertigten und heiligten“.103
In Schulen und Hochschulen damals wurden Affen, „minderwertige“ Rassen und „nordische Edelmenschen“ in einer Reihe der Höherentwicklung als Tatbestände angeboten. Nur der Stärkere und Fähigere war zu Hochkulturen befähigt. Der Minderwertige, Schwache und Kranke aber muss ausgemerzt werden, weil er den Stärkeren und Gesunden Lebensraum wegnimmt. So erklärt sich die Euthanasie von Behinderten und Kranken und die Beseitigung unerwünschter Rassen, die zu Untermenschen oder gar Unmenschen gestempelt wurden. Diese Rassentheorie wurde von der Molekularbiologie widerlegt, da die Analyse der Mitochondrien-DNA zeigen konnte, dass die Menschen aller Kontinente engstens miteinander verwandt sind und der zoologische Begriff „Rasse“ keinerlei Berechtigung hat.
Die Auslegungen einer naturwissenschaftlichen Theorie sind sehr bedauerlich. Hier könnte Aufklärung hilfreich sein.
Schluss: Welche Einstellung empfiehlt das Berufsethos dem Naturwissenschaftler zur Evolutionslehre?
Vier verschiedene Vorstellungen bemühen sich, die Probleme der Entstehung des Lebens und der Vielfalt der Organismen zu lösen:
1. Die herrschende Evolutionstheorie baut „nach Mäßstäben von Atomen und Molekülen“ (Crick) auf Darwins Selektion umweltverträglicher Varianten der Arten durch natürliche Zuchtwahl auf, der Mikroevolution. Nach dieser Selektionstheorie zufällig entstandener Varianten entstehen unter den gegebenen Notwendigkeiten der Umwelt i. A. schrittweise sowohl auf der molekularen Ebene das Leben als auch auf der Ebene der Organismen immer höher entwickelte Lebewesen mit immer komplexeren Einrichtungen, wie Auge, Gehirn oder auch der Geißelmotor der Bakterien. Manche Anhänger dieser Theorie meinen, der Übergang von der Mikroevolution zur Makroevolution sei kontinuierlich.104
2. Die theistische Evolutionstheorie, nach der die Entstehung des Lebens und die Höherentwicklung der Organismen nicht nur auf Materie und Energie beruht, sondern dass dazu das geistige Prinzip der Information erforderlich ist, die von einem planenden Schöpfer stammt.
3. Der Nachweis des Schöpfers, des geistigen Verursachers, der von Albert Einstein als „unendlich überlegene Vernunft“ bezeichnet wurde, wird als Intelligent Design-Konzept besonders von Naturwissenschaftlern in den USA diskutiert. Nach diesem sollen die Signale für Entstehung, Prozesse und Bau von Komplexitäten des Lebens erforscht werden.
4. Die Schöpfungswissenschaft basiert auf der in den Bibelberichten enthaltenen inspirierten Wahrheit, die auch über die Entstehung der Organismenvielfalt Auskunft und Impulse für die Forschung gibt. Hierbei ist die korrekte Übersetzung der Bibelberichte von großer Bedeutung. Gott schuf die Organismen als Einheiten der Schöpfung „nach ihren Arten“105. Diese Schöpfungseinheiten bilden Grundtypen, die sich durch ihren Habitus und genetisch feststellbare große Ähnlichkeit auszeichnen, wobei die Abgrenzung der Einheiten durch die Taxa der Systematik von Gattungen bis zu den Familien, selbst bei Bakterien, gegeben ist. Eine Evolution findet nur innerhalb der Grundtypen statt, Übergänge zwischen den Grundtypen sind nicht möglich und nicht bekannt.
Nobelpreisträger Sir John Eccles äußerte einmal zur Frage der Makroevolution: „… das wissenschaftliche Establishment ist doch total wider mich. Selbst mit Nobelpreis und 17 Ehrendoktoraten bin ich total ausgegrenzt, einsam. Würde ich noch die Entwicklungsgeschichte des Gehirns in Frage stellen, würde nichts mehr von dem Erreichten diskutiert werden.“
Dies zeigt, mit welcher Radikalität die Evolutionstheorie als Ideologie eines „Evolutionismus“ in manchen Institutionen vorgetragen wird, anstatt sie sachlich anzugehen. Mit dem Nachweis der Variabilität begann Darwin und fand die Selektion als Mittel zur Entstehung neuer Formen, wie sie Pflanzen- und Tierzüchtung unter der Hand des Menschen kennt und nun von Darwin auf die natürliche Zuchtwahl angewendet und dadurch zu einer fruchtbaren Theorie wurde, die zahlreiche Forschungsprogramme initiierte. Doch in dem Maße wachsender Schwierigkeiten mit neuen Erkenntnissen wurde die Evolution (Makroevolution) von Ideologen zu einer „bewiesenen Wissenschaft“ erhoben. So stellt der Biochemiker Herbert Niellson fest, die Evolution sei ein schweres Hindernis“ für die biologische Forschung; denn sie „verhindert nämlich konsequente Schlussfolgerungen aus sogar dem Experimentalmaterial… Alles muss letzen Endes nach dieser spekulativen Theorie gebogen werden... Eine exakte Biologie kann deshalb nicht aufwachsen“106. Der Mikrobiologe Stefan Bleeken schreibt: „Die Biologie befindet sich in einem zur Physik vor Newton vergleichbaren Entwicklungsstand und ihr Weg zu einer formalen Wissenschaft liegt im Dunkeln… Die Molekularbiologie hat ihren Ehrgeiz aufgegeben, das Funktionieren eines lebenden Organismus zu erklären und ihre Forschungsstrategie wegen des Fehlens eines theoretischen Rahmens ist zur reinen Daten- und Faktensammlung verflacht“.107 D. W. S. Watson fand die Begründung für die Radikalität der Ideologen: „Die Evolution selbst wird akzeptiert, nicht weil man Derartiges beobachtet hätte, oder weil man sie durch eine logisch zusammenhängende Beweiskette als richtig beweisen könnte, sondern weil die einzige Alternative dazu – der Schöpfungsakt Gottes – einfach unmöglich ist.“ Angesichts der Zweifel stellt sich die Frage, welche Einstellung dem korrekten Naturforscher angemessen ist.
Es sei bei dieser Frage nochmals klargestellt: In der Makroevolution wurden weder für eine echte Höherentwicklung noch für die Entstehung des Lebens, der Entstehung von Information aus Nichtinformation, durch Zufall und Versuch und Irrtum, „trial and error“, überzeugende Anhaltspunkte gefunden, wohl aber für den umgekehrten Weg der Degeneration, des Verfalls, bei der durch Zerstörung eines komplexen Mechanismus eine Anpassung an widrige Lebensverhältnisse gefunden werden kann, das aber sich unmöglich für die Argumentation einer Anpassung im Sinne einer Höherentwicklung benutzen lässt. Die Forschungsergebnisse der Mikroevolution, der Variation der Arten, haben seit alters in der Tier- und Pflanzenzüchtung bereits eine experimentelle Grundlage gehabt und bilden Darwins eigentliches Werk als Frucht einer systematischen Arbeit. In den Streitgesprächen über die Evolutionstheorie argumentieren ihre radikalen Anhänger mit Beweisen aus der Mikroevolution, während die Kritiker der Evolutionstheorie stets auf den Beweisnotstand der Makroevolution weisen und die andersartige Mikroevolution als bewiesen ansehen.
Die einleitende Feststellung der Zeitschrift „Nature“, (approved by the University board of Regents (2006)), mahnt in Fortführung des Satzes „Evolution by natural selection is a theory, not a fact. This material should be approached with an open mind, studied carefully and critically considered“. Dem ist nichts hinzuzufügen.
Weiterführende Literatur
- Arber W., Spontanmutation, Mechanismen der biologischen Evolution, Naturw Rdsch (1997); 50: 1-9
- Bürgi L., Geheimakte Archäologie, Herbig-Verlag, Bettendorf (1998)
- Gitt W., Das sonderbarste Schiff der Weltgeschichte, Sonderdruck aus Fundamentum 3/2000, Immanuel-Verlag, Ravensburg (2002), 2. Auflage
- Herder, der neue, von A bis Z, Herder Verlag, Freiburg (1949), 9. Auflage
- Johnson P. E., Darwin im Kreuzverhör, CLV-Christliche Literaturverbreitung, Bielefeld (2003)
- Johnson P. E., Reason in the Balance, InterVarsity Press, Downers Grove (1998)
- Junker R., Die Frage nach dem Urheber, factum (2005); 7: 28-37
- Junker R., Unerhört: Konvergenz eines Schlüsselmerkmals, Studium Integrale Journal (2005); 12: 74-76
- Kang, C. H., Nelson E. R., Erinnerungen an die Genesis – Die Chinesen und die biblische Urgeschichte, Hänssler-Verlag Holzgerlingen (1998)
- Rensch B., Neuere Probleme der Abstammungslehre – die transspezifische Evolution, Encke Verlag, Stuttgart (1947)
- Spaemann R., Freiheit der Forschung oder Schutz des Embryos?, Die Zeit, 20. 11. 2003 (Nr. 48)
Referenzen
- Spencer H., „Kampf ums Dasein“, „Überleben der Tüchtigsten“, in: Jahn I., Geschichte der Biologie, Nikol Verlag, Hamburg (2004), 3. Auflage, S. 960
- Kutschera U., Evolutionsbiologie. Eine allgemeine Einführung, Parey Buchverlag, Berlin (2001)
Kämpfe L. (Hrsg), Evolution und Stammesgeschichte der Organismen, Gustav-Fischer-Verlag, Stuttgart (1980)
Schick R. (Hrsg), Evolution, Gustav Fischer-Verlag, Stuttgart (1987), 3. Auflage
Grassé P. P., Evolution, Allgemeine Biologie (Bd. 5), Fischer Verlag, Stuttgart (1973)
Junker R., Scherer S., Evolution, ein kritisches Lehrbuch, Weyel-Lehrmittelverlag, Gießen (2006), 6. Auflage
Beck H. W., Variationen zu einer interdisziplinären Schöpfungskosmologie, Peter Lang Verlag, Frankfurt (1999)
Behe M. J., Darwins Black Box, Michael Behe (1996)
Gitt W., Auslegungssätze zur Bibel aus der Sicht eines Naturwissenschaftlers (2. Teil), Theol. Beilage zur STH-Pastille Nr. 3, 2006
Grün J., Die Schöpfung, ein göttlicher Plan, Verax-Verlag Müstair (2000)
vom Stein A., Creatio, Biblische Schöpfungslehre, Daniel-Verlag, Lychen (2005) - Scheven J., Daten zur Evolutionslehre im Biologieunterricht, Hänssler-Verlag, Neuhausen/Stuttgart (1979)
- Hartmann M., Die philosophischen Grundlagen der Naturwissenschaften, Gustav Fischer Verlag, Stuttgart (1959), 2. Auflage
- Höfling O., Physik, Ferdinand Dümmler Verlag, Berlin (1994), 15. Auflage
- Geflügeltes Wort aus der Arbeitsgruppe des Nobelpreisträgers Butenandt. Der Autor hatte bei einem direkten Schüler von Butenandt promoviert, so dass er dieses Zitat sozusagen als persönliche Mitteilung übernommen hat.
- Portmann A., Entläßt die Natur den Menschen?, Piper Verlag, München (1970)
- Zentrale Quelle für die „Vorgeschichte der Lehre Darwins“ ist das sehr umfangreiche Buch von Ilse Jahn (Hrsg), Geschichte der Biologie, Nikol Verlagsgesellschaft Hamburg (2004), 3. Auflage
- Brandt M., Wie alt ist die Menschheit?, Hänssler-Verlag Holzgerlingen (2006)
- Jahn I. (Hrsg), Geschichte der Biologie, Nikol Verlagsgesellschaft Hamburg (2004), 3. Auflage
- Palfy J., Katastrophen der Erdgeschichte, Schweizer-bart’sche Verlagsbuchhandlung, Stuttgart (2005)
- Junker R., Scherer S., Evolution, ein kritisches Lehrbuch, Weyel-Lehrmittelverlag, Gießen (2006), 6. Auflage
- Jahn I. (Hrsg), Geschichte der Biologie, Nikol Verlagsgesellschaft Hamburg (2004), 3. Auflage
- Junker R., Zur Abgrenzung von Mikroevolution und Makroevolution, Studium Integrale Journal (2006); 13: 59-67
- Kowalevski W., Aus der Monographie der Gattung Anthropotherium von Cuvier und Versuch einer natürlichen Classifikation der fossilen Huftiere, in: Palaeontographica, Beiträge zur Naturgeschichte der Vorwelt, S. 131-346
- Rich T. H. et al., Independent Origins of Middle Ear Bones in Monotremes and Therians, Science (2005); 307: 910-914
- vgl. Jahn I. (Hrsg), Geschichte der Biologie, Nikol Verlagsgesellschaft Hamburg (2004), 3. Auflage
- Blechschmidt E., Wie beginnt das menschliche Leben? Vom Ei zum Embryo, Christiana-Verlag, Stein am Rhein (2002)
- vgl. Jahn I. (Hrsg), Geschichte der Biologie, Nikol Verlagsgesellschaft Hamburg (2004), 3. Auflage
- Jahn I. (Hrsg), Geschichte der Biologie, Nikol Verlagsgesellschaft Hamburg (2004), 3. Auflage
- vgl. Darwin C., Die Entstehung der Arten durch natürliche Zuchtwahl, Reclam Verlag, Leipzig (1963)
- vgl. Jahn I. (Hrsg), Geschichte der Biologie, Nikol Verlagsgesellschaft Hamburg (2004), 3. Auflage
- vgl. Jahn I. (Hrsg), Geschichte der Biologie, Nikol Verlagsgesellschaft Hamburg (2004), 3. Auflage
- Jahn I. (Hrsg), Geschichte der Biologie, Nikol Verlagsgesellschaft Hamburg (2004), 3. Auflage
- Rich T. H. et al., Independent Origins of Middle Ear Bones in Monotremes and Therians, Science (2005); 307: 910-914
- Dombrowski H. J., Bacillus circulans aus Zechsteinsalzen, Zentralalblatt Bakteriol (1960); 183: 173-179
- Vogel G., Tracking the History of the genetic code, Science (1998); 281: 329-331
- Randerson J. P., Hurst L. D., The uncertain evolution of the sexes, Trends Ecol Evolution (2001); 16: 571-579
- Schmitt R., Molekulare Propeller: Bakteriengeißel und ihr Antrieb, Biologie in unserer Zeit (1997); 27: 40-47
- Portmann A., Entläßt die Natur den Menschen?, Piper Verlag, München (1970)
- Vollmert B., Das Molekül und das Leben, Rowohlt Verlag, Reinbek (1985)
- Eigen M., Stufe zum Leben, Piper Verlag, München (1992), 2. Auflage
- Junker R., Scherer S., Evolution, ein kritisches Lehrbuch, Weyel-Lehrmittelverlag, Gießen (2006), 6. Auflage
- zit. n. Gitt W., Am Anfang war die Information, Hänssler-Verlag, Holzgerlingen (2002), 3. Auflage
- Eigen M., Stufe zum Leben, Piper Verlag, München (1992), 2. Auflage
- Kuhn W., Keine Schöpfung ohne Schöpfer, Wissenschaftliche Bildung und Glaube, Wachenheim Verlag (ohne Jahresangabe)
- Husak P., Gibt es eine akzeptable Deszendenztheorie für den Menschen? Kritische Reflexionen über die Phylogenese und Ontogenese des Menschen, Dissertation an der Intern. Akad. für Philosophie im Fürstentum Liechtenstein (2004)
- Kutschera U., Evolutionsbiologie. Eine allgemeine Einführung, Parey Buchverlag, Berlin (2001)
- Hartwig-Scherer S., Die Zwergmenschen – Genetische Isolation führt zur Bildung von „Hobbits“ auf indonesischer Insel, Studium Integrale (2005); 12(1)
- Cremo M. A., Thompson R. L., Verbotene Archäologie, Sensationelle Funde verändern die Welt, Bechtermünz-Verlag, Augsburg (1996)
Liebi R., Herkunft und Entwicklung der Sprachen, Hänssler-Verlag, Holzgerlingen (2004), 2. Auflage
Zillmer H.-J., Darwins Irrtum, Langen Müller, Herbig-Verlag, München (2006), 8. Auflage - Junker R., Scherer S., Evolution, ein kritisches Lehrbuch, Weyel-Lehrmittelverlag, Gießen (2006), 6. Auflage
- Schrenk J., Die Frühzeit des Menschen, C. H. Beck Verlag, München (1997)
- Sadler T. W., Medizinische Embryologie, Thieme Verlag, Stuttgart (1998)
Süßmuth R., Süßmuth S., Die Befruchtung: Die Bildung der Zygote – der Anfang des Menschen, Medizin und Ideologie (2003); 25: 11-14
Beckmann R., Löhr M. (Hrsg.), Der Status des Embryos, Johann Wilhelm Naumann Verlag, Würzburg (2003) - Eccles J. C., Wie das Selbst sein Gehirn steuert, Piper Verlag, München (1994)
- Thürkauf M., Die moderne Naturwissenschaft und ihre soziale Heilslehre – der Marxismus, Novalis-Verlag, Mannheim (1980)
- Thürkauf M., Die moderne Naturwissenschaft und ihre soziale Heilslehre – der Marxismus, Novalis-Verlag, Mannheim (1980)
- Junker R., Scherer S., Evolution, ein kritisches Lehrbuch, Weyel-Lehrmittelverlag, Gießen (2006), 6. Auflage
- Blechschmidt E., Wie beginnt das menschliche Leben? Vom Ei zum Embryo, Christiana-Verlag, Stein am Rhein (2002)
- Portmann A., Entläßt die Natur den Menschen?, Piper Verlag, München (1970)
- Portmann A., Entläßt die Natur den Menschen?, Piper Verlag, München (1970)
- Lindner E., Wahrheit und Wirklichkeit, M. Lindner-Verlag, Karlsruhe (1997)
- Behe M. J., Darwins Black Box, Michael Behe (1996)
- Junker R., Scherer S., Evolution, ein kritisches Lehrbuch, Weyel-Lehrmittelverlag, Gießen (2006), 6. Auflage
- Johnson P. E., Darwin on trial, InterVarsity Press, Downers Groven (1993), 2. Auflage
- Ruse M., Is Evolution a Secular Religion?, Science (2003); 299: 1523-1524
- Schmitt R., Molekulare Propeller: Bakteriengeißel und ihr Antrieb, Biologie in unserer Zeit (1997); 27: 40-47
- Gitt W., Am Anfang war die Information, Hänssler-Verlag, Holzgerlingen (2002), 3. Auflage
- Grzimek B., Vögel 3, in: Grzimeks Tierleben, Deutscher Taschenbuch-Verlag, München (1979), S. 483
- Kayser F. et al., Medizinische Mikrobiologie, Thieme-Verlag, Stuttgart (1998), 9. Auflage
- Junker R., Scherer S., Evolution, ein kritisches Lehrbuch, Weyel-Lehrmittelverlag, Gießen (2006), 6. Auflage
- Gen 1, 28
- Jer 1, 5
- Ps 139, 15 ff
- Weish 8, 11, I. Vatik. K., DS 3003
- Ps 33 (32), 9
- Gen 1, 3 ff
- Husak P., Gibt es eine akzeptable Deszendenztheorie für den Menschen? Kritische Reflexionen über die Phylogenese und Ontogenese des Menschen, Dissertation an der Intern. Akad. für Philosophie im Fürstentum Liechtenstein (2004)
- Scheven J., pers. Mitteilung 2006 – nach Altermann (1999) und Klemm (2000), Kuratorium Lebendige Vorwelt e. V., D-65711 Hofheim
- Gen 11, 11-21
- Junker R., Scherer S., Evolution, ein kritisches Lehrbuch, Weyel-Lehrmittelverlag, Gießen (2006), 6. Auflage
- Stephan M., Fritsche T., Sintflut und Geologie, Hänssler-Verlag, Holzgerlingen (2002)
- Dürr H. D., Elemente des Lebens, SFG Service Center Fachverlag, Kusterdingen (2000)
- Stephan M., Fritsche T., Sintflut und Geologie, Hänssler-Verlag, Holzgerlingen (2002)
- zit. nach Zillmer H.-J., Irrtümer der Erdgeschichte, Herbig-Verlag, München (2003)
- Stephan M., Fritsche T., Sintflut und Geologie, Hänssler-Verlag, Holzgerlingen (2002)
- Stephan M., Fritsche T., Sintflut und Geologie, Hänssler-Verlag, Holzgerlingen (2002)
- Norman T., Setterfield B., The Atomic Constants: Light and Time, SRI International (1987)
- Walter K.: Schneller als das Licht, Faktum (2003); 23(5): 29-31
- Brandt M., Wie alt ist die Menschheit?, Hänssler-Verlag, Holzgerlingen (2006)
- Gen 10, 2, ferner 1 Chr 1, 19
- Gitt W., Am Anfang war die Information, Hänssler-Verlag, Holzgerlingen (2002), 3. Auflage
- Scheven J., Vor uns die Sintflut. Eine Kritik der aktualistischen Geologie, Kuratorium Lebendige Vorwelt, Hofheim a. T. (2007)
- Ez 31, 18
- Scheven J., Karbonstudien, Wort und Wissen Bd. 18, Hänssler Verlag, Neuhausen/Stuttgart (1986)
- Scheven J., Mega-Sukzessionen und Klimax im Tertiär, Wort und Wissen Bd, 19, Hänssler Verlag, Neuhausen/Stuttgart (1988)
- Scheven J., pers. Mitteilung 2005
- Stephan M., Fritsche T., Sintflut und Geologie, Hänssler-Verlag, Holzgerlingen (2002)
- Der Begriff „Wolfskapitalismus“ wird Peter Scholl-Latour zugeschrieben.
- „Was anderes sind Reiche, wenn Gerechtigkeit fehlt, als große Räuberbanden.“; Augustinus, Vom Gottesstaat, in: Augustinus, Bücher 1-22, Bd. I, Buch 1-10, Buch 4, Düsseldorf-Zürich (1997), 4. Auflage, S. 173
- Uschmann G., Nachwort, in: Darwin C., Die Entstehung der Arten durch natürliche Zuchtwahl mit Nachwort von Georg Uschmann, Reclam Verlag, Leipzig (1949)
- Uschmann G., Nachwort, in: Darwin C., Die Entstehung der Arten durch natürliche Zuchtwahl mit Nachwort von Georg Uschmann, Reclam Verlag, Leipzig (1949)
- Klaus G., Buhr M. (Hrsg.), Philosophisches Wörterbuch, VEB Bibliographisches Institut, Leipzig (1976)
- Klaus G., Buhr M. (Hrsg.), Philosophisches Wörterbuch, VEB Bibliographisches Institut, Leipzig (1976)
- Grün J., Die Schöpfung, ein göttlicher Plan, Verax-Verlag, Müstair/GR (2000)
- Schwemmler P., Kosmos, Erde, Mensch. Vom wahren Wesen der Evolution, Kohlhammer Verlag, Stuttgart (1997)
- Bergmann, in: Gitt W., Am Anfang war die Information, Hänssler-Verlag, Holzgerlingen (2002), 3. Auflage
- Junker R., Leben durch Sterben? Schöpfung, Heilsgeschichte und Evolution, Hänssler-Verlag, Neuhausen/Stuttgart (1994), 2. Auflage
- Morris D., Das Tier Mensch, eine Naturgeschichte des Homo sapiens, Video-Aufzeichnung (Timelife Video), BBC Worldwide Ltd. (1995)
- Blechschmidt E., Wie beginnt das menschliche Leben? Vom Ei zum Embryo, Christiana-Verlag, Stein am Rhein (2002)
- Auer A., in: Rhonheimer M., Natur als Grundlage der Moral, Tyrolia Verlag, Innsbruck (1987), S. 149
- Drewermann E., … und es geschah so, Walter-Verlag, Zürich, Düsseldorf (1999)
- Anatomie des SS-Staates, Bd. 2, dtv-Dokumente 2916, S. 246, zit. nach Grün J., Die Schöpfung, ein göttlicher Plan, Verax-Verlag, Müstair/GR (2000)
- Fromm E., Anatomie der menschlichen Destruktivität, Rohwolt Taschenbuch, Hamburg (1977), S. 49, zit. nach Grün J., Die Schöpfung, ein göttlicher Plan, Verax-Verlag, Müstair/GR (2000)
- Junker R., Zur Abgrenzung von Mikroevolution und Makroevolution, Studium Integrale Journal (2006); 13: 59-67
- Gen 1, 21; 1, 24; 1, 25
- Gitt W., Am Anfang war die Information, Hänssler-Verlag, Holzgerlingen (2002), 3. Auflage
- Gitt W., Am Anfang war die Information, Hänssler-Verlag, Holzgerlingen (2002), 3. Auflage
Prof. Dr. rer. nat. Roland Süßmuth
Am Buchenhain 18, D-72622 Nürtingen
EvonOstrowski(at)web.de
Gustav-Siewerth-Akademie
D-79809 Weilheim-Bierbronnen



