Von der Entwicklungs- zur Evolutionstheorie
Zusammenfassung
Evo-Devo („Evolutionary developmental biology“) ist ein neuer Wissenschaftszweig, der vielversprechende Einsichten innerhalb der Evolution als auch der Entwicklungsbiologie erlaubt. Die Idee, diese beiden Forschungsfelder zu vereinen, ist allerdings nicht neu. Dieser Artikel beschreibt zunächst die Geschichte der Verbindung zwischen der Entwicklungsbiologie und Evolution, also quasi die Evolution von Evo-Devo. Danach werden die Fragestellungen der modernen Evo-Devo vorgestellt und anhand von Beispielen gezeigt, wie Evo-Devo weiteres Licht auf die Prozesse wirft, die zur Evolution der Arten führen können, nämlich durch die Abänderung der Regulationsprozesse während der Entwicklung. Dieses Wissen kann und wird die klassische Evolutionstheorie erweitern.
Schlüsselwörter: Entwicklungsbiologie, Evolution, Evo-Devo
Abstract
Evo-Devo („Evolutionary developmental biology“), a new branch of science, shows promising insights into our understanding of evolution and developmental biology. But the idea of bringing together these two fields is not a new one. This article describes on the one hand the history of interactions between developmental biology and evolution, virtual explaining the evolution of Evo-Devo. Later the research interests of modern Evo-Devo are explained and some examples are shown of how this field is able to shed some light on how species evolve, namely by changing regulatory patterns during development. This knowledge can and will expand the classical theory of evolution.
Keywords: Development, Evolution, Evo-Devo
Entwicklungs- und Evolutionsbiologie – zwei voneinander unabhängige Fachdisziplinen?
Zunächst erscheint es, als wären die Entwicklungs- und die Evolutionsbiologie zwei zwar verwandte, aber dennoch unterschiedliche Wissenschaftsbereiche. Der größte Unterschied liegt in der Fragestellung. Im Zentrum der entwicklungsbiologischen Forschung liegt die unveränderte Weitergabe des Körperbauplans von den Eltern an die Nachkommen, oder anderes formuliert: Wie entsteht aus einer befruchteten Eizelle ein Adulttier, das genau wie seine Elterntiere aussieht? Dasselbe gilt im Übrigen auch für die Pflanzen. Die Evolutionsbiologie hingegen ist genau an diesen Abänderungen im Körperbauplan interessiert. Als kleines Beispiel ist das Pferd geeignet. Aus dem kleinen Urpferd entstand durch kleine Mutationen und Selektionen ein großes Tier, das zum Tragen von Lasten und Ziehen von Wägen geeignet war. Durch den Menschen erfolgte dann eine gezielte Züchtung (d. h. wieder Selektion) zu einer Form, die befähigt war, auch Reiter zu tragen.1 Bei diesem Beispiel geht es um vergleichsweise geringe Abänderungen, wenn man die Unterschiede zwischen einem einzelligen Lebewesen und einem Wirbeltier bedenkt, z. B. dem Menschen. (Abb. 1)
Wie oder besser gesagt, wann entsteht aber die Veränderung? Es ist ja nicht so, dass ein erwachsenes Tier oder eine Pflanze plötzlich eine neue Eigenschaft entwickelt. Es sind immer die Nachkommen, die durch Mutation und Rekombination ihres genetischen Materials sich von ihren Eltern unterscheiden. Im vorher genannten Beispiel wäre das eine verstärkte Wirbelsäule und Muskulatur des Pferdes. François Jacob (1977) formulierte dieses so: „Die Evolution arbeitet mit dem, was sie hat. Sie kombiniert existierende Teile auf neue Arten miteinander. Dies passiert auf der Ebene der Gene, die die Entwicklung steuern und nicht der der Adultgene.“2
Natürlich gibt es auch hier einige Ausnahmen. So können zum Beispiel Bakterien und Viren genetisches Material an andere Bakterien oder befallene Lebewesen weitergeben und diese somit verändern. Antibiotikaresistenzen werden auf diese Art und Weise zwischen Bakterien ausgetauscht.3
Eine weitere Ausnahme könnte bei der Entstehung der Eukaryonten vorliegen. In jeder eukaryontischen Zelle, d. h. Zellen mit echtem Zellkern, gibt es Mitochondrien und in pflanzlichen auch noch Chloroplasten. Die Mitochondrien sind verantwortlich für den Energiestoffwechsel und die Chloroplasten für die Photosynthese. Diese beiden Arten von Zellorganellen besitzen eigene DNA. Diese zeigt nun in genetischen Vergleichen Eigenschaften von prokaryontischer DNA. Man nimmt daher an, dass prokaryontische Zellen in die Vorform der eukaryontischen Zellen eingewandert sind und dort nicht verdaut wurden, sondern eine Symbiose mit der Wirtszelle eingegangen sind. Diese Theorie wird als Endosymbiontheorie bezeichnet.4
Ein weiterer Berührungspunkt der beiden Disziplinen kann gefunden werden, wenn man bedenkt, dass während der Evolution aus einem einzelligen Lebewesen höher entwickelte vielzellige Arten entstanden sind und während der Entwicklung ebenfalls aus einem Einzeller (hier die befruchtete Eizelle) viele spezialisierte Zellen, die Organe bilden, ein komplexer Organismus aufgebaut wird. Tab. 1 fasst diesen Vergleich noch einmal zusammen.
| Evolution | Entwicklung |
|---|---|
| Änderung im Körperbauplan | Weitergabe des unveränderten Körperbauplans von den Eltern an die Nachkommen |
| Evolution beinhaltet auch die Änderungen in der Embryonalentwicklung | |
| Aus einfachen Lebewesen (Einzeller) entwickeln sich höhere Lebewesen (z. B. Wirbeltiere) | Eine Zelle entwickelt sich in verschiedene Arten von spezialisierten Zellen (Differenzierung) |
Tab. 1: Vergleich zwischen Evolution und Entwicklung
Historische Verknüpfungsversuche
Das Interesse, die Evolutions- mit Entwicklungsbiologie zu verknüpfen, ist, seitdem es beide Disziplinen gibt, vorhanden. Ernst Haeckel und Karl Ernst von Baer sind wohl die berühmtesten Wissenschaftler, die dieses versucht haben.
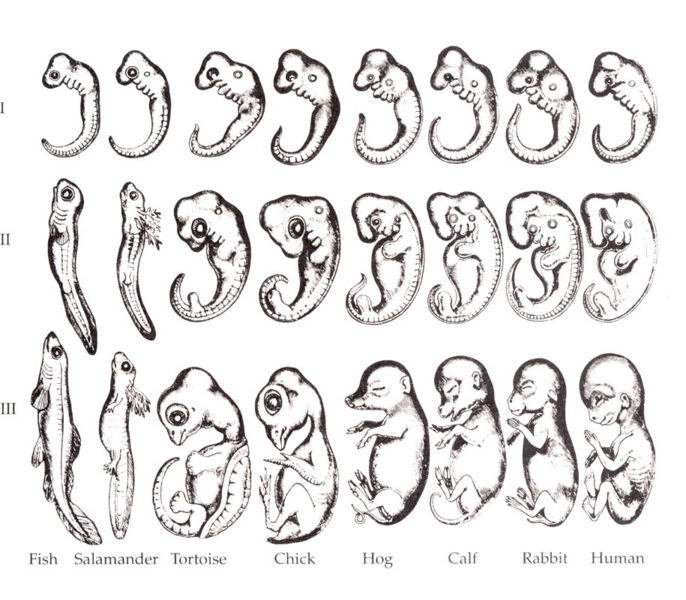
Ernst Haeckel versuchte es mit der Formulierung des Biogenetischen Gesetzes, welches besagt, dass die Ontogenese die Phylogenese wiederholt. Haeckels Hypothese lautete, dass jedes Lebewesen während seiner Entwicklung frühere Stadien der Evolution durchläuft (Abb. 2).5 So soll zum Beispiel jedes Säugetier zunächst fisch- und reptilartige Formen durchlaufen haben, bevor es die säugetierartigen Charakteristika zeigt. Das Biogenetische Gesetz war die Folge der fälschlichen Annahme, die Evolution sei ein linearer Prozess, bei dem aus einfacheren Lebewesen immer komplexere entstehen. Erwähnungswert ist hier, dass Haeckel für die Gegenüberstellung der verschiedenen Embryonen (Abb. 2) die Zeichnungen für die Tafeln so anfertigte, dass Ähnlichkeiten übertrieben dargestellt wurden und Unterschiede nicht abgebildet wurden. Kurz gesagt, er veränderte sie so, dass sie in seine Theorie passten.
Die heute anerkannte Evolutionstheorie geht hingegen davon aus, dass sich die einzelnen Arten durch Veränderung von ihren Vorformen abgespalten haben, also nicht linear, sondern verzweigt.
Interessanter Weise nutzte Haeckel für die Formulierung seines Gesetzes die Baer’schen Regeln oder das Gesetz der Embryonenähnlichkeit, das eben nicht davon ausgeht, dass ein Embryo frühere Stadien durchläuft.
Die Regeln beruhen auf vier Grundsätzen:
- Embryonen verschiedener Arten sind umso schwerer voneinander zu unterscheiden, je jünger sie angetroffen werden.
- Weniger allgemeine Merkmale entwickeln sich aus den allgemeineren, bis schließlich die spezialisierten erscheinen.
- Der Embryo einer Spezies entwickelt sich nicht, indem er die Stadien niedriger Stufen durchläuft, sondern indem er sich mehr und mehr von diesen entfernt.
- Aus diesem Grunde ist der Embryo eines höher entwickelten Embryos niemals wie ein niederes Tier, sondern nur wie dessen früherer Embryo.
Anknüpfungsversuche im 20. Jahrhundert – Die synthetische Evolutionstheorie
Im Jahre 1922 lehnte Walter Garstrang die vorher genannten Verknüpfungsversuche ab, indem er erklärte, dass die Ontogenie nicht die Phylogenie kopiert, sondern dass die Ontogenie die Grundlage der Phylogenie sei. „Der erste Vogel schlüpfte aus einem Reptilei“, so brachte Garstrang es auf den Punkt. Damit meinte er, wie schon eingangs erwähnt, dass im Bauplan der Nachkommen von Reptilien Vögel durch graduelle Veränderungen entstanden. Aber genau dieses langsame Fortschreiten ist ein Problem dieser Theorie. Wie sollen so neue Strukturen entstanden sein? Ein Beispiel: Reptilien haben Schuppen und Vögel haben Federn. Kleinere genetische Veränderungen in den Proteinen, die diese Strukturen aufbauen, würden sehr wahrscheinlich zu nicht funktionellen Zwischenprodukten führen, die dadurch ausselektiert werden würden. Dieses ist übrigens genau der Punkt, an dem Vertreter der Intelligent Design und Scientific Creationism Theorie ansetzen.
Richard Goldschmidt (1940) konnte die Aussage Garstrangs verfeinern, indem er erklärte: „Die Ansammlung kleiner genetischer Veränderungen sind nicht ausreichend, um evolutionäre neue Strukturen wie z. B. die Zähne, Federn oder Schildkröten-Panzer zu erschaffen. Diese Änderungen entstehen durch Mutationen in Genen, die die Entwicklung regulieren.“6 Der Unterschied liegt also darin, dass nicht die strukturellen Gene verändert werden, also die Gene, die für den Aufbau von Organen zuständig sind, sondern die Gene, die für die Regulation des Entwicklungsprozess wichtig sind.
Ein Beispiel hierfür ist die unterschiedliche Entwicklungssteuerung für Hühner- und Entenfüße (Abb. 3).7 Der Unterschied ist klar erkennbar: Ein Hühnerfuß hat Krallen und kleine Schuppen bedecken ihn, bestens geeignet, um im Sand zu scharren. Die Ente auf der anderen Seite braucht ihre Füße zum Paddeln im Wasser und ist mit ihren Schwimmhäuten perfekt dafür ausgerüstet. Die Grundentwicklung beider Fußarten verläuft gleich. Um aber feingliedrige Zehen oder aber auch Finger zu bilden, muss das überschüssige Gewebe durch Zelltod zerstört werden. Ein Gen, das diesen aktiviert, ist Bmp. In Abbildung 3 sieht man, dass sowohl im Huhn als auch in der Ente dieses Gen aktiv ist. Um nun im Entenfuß zu verhindern, dass auch die zukünftigen Schwimmhäute abgebaut werden, wird Gremlin, das die Aktivität des Proteins BMP hemmt, auch in dem Bereich gebildet, wo später die Schwimmhäute sein werden und nicht nur im Zehenbereich wie beim Huhn. Es hat bei dieser Änderung also das Regulationsgen Gremlin seinen Wirkungsort geändert.
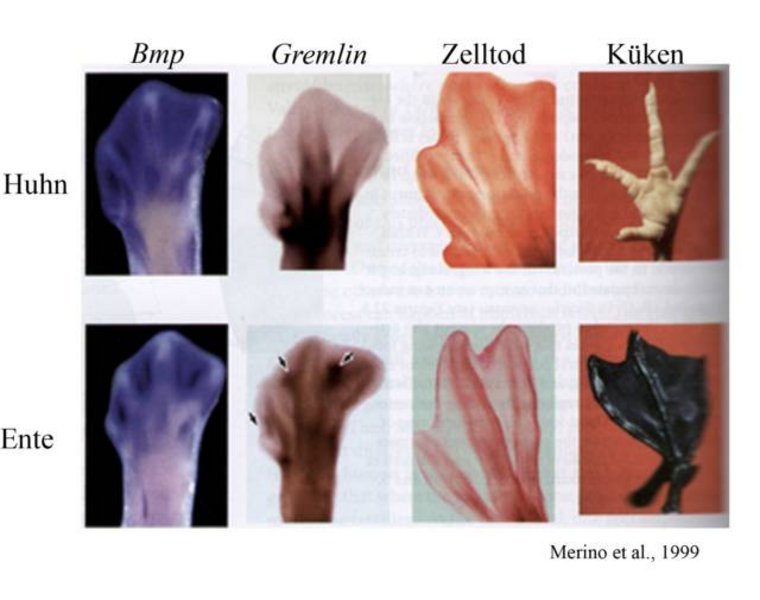
Dieses Beispiel zeigt auch, wie sich aus der synthetischen Evolutionstheorie, das neue Feld „Evolutionäre Entwicklungsbiologie“ oder umgangssprachlich Evo-Devo entwickelt hat. Wichtig für diese Entwicklung waren Werke wie J. Goulds „Ontogeny and Phylogeny“8 und „Evolution by tinkering“ von dem bereits erwähnten Francois Jacob.9 Aber der große Durchbruch von Evo-Devo kam um das Jahr 2000. Der Grund lag in der Etablierung von molekularen Techniken wie z. B. dem Sequenzieren, PCR usw. in beiden Feldern. Die Evolutionäre Entwicklungsbiologie beschäftigt sich mit drei Schwerpunkten:
(1) Welche Ähnlichkeiten können in der Regulation von Entwicklungsprozessen in verschiedenen Arten gefunden werden?
(2) Was sind die Modifikationen dieser Regulationsmechanismen? Hier spielen die genetischen Unterschiede der Regulationsgene, aber auch die Expression gleicher Gene zu unterschiedlichen Zeiten und Stellen im Embryo, wie z. B. im vorhin erwähnten Beispiel von der Fußentwicklung bei Huhn und Ente, eine Rolle.
(3) Wie variieren diese Gene und ihre Expression innerhalb einer Population und können so neue Arten entstehen?
Als Beispiel für den ersten Schwerpunkt werden gerne die Hoxgene verwendet. Mitglieder dieser Genfamilie kommen im gesamten Tierreich vor. In allen Tieren sind sie wichtig für die Polarität des Tieres, wo Kopf und Schwanz ist, aber auch wo Beine gebildet werden, und bei Insekten, wo Antennen wachsen sollen. Ein weiteres interessantes Beispiel für die Ähnlichkeit von Regulationsprozessen ist die Augenentwicklung. Obwohl durch das Zusammenspiel verschiedener Regulationsgene, die vielfältigen Augenarten gebildet werden, wie z.B. unser Linsenauge, oder das Facettenauge der Insekten, gibt es ein Gen, dass in allen Tieren die Augenentwicklung „anschaltet“: das Gen Pax6. Walter Gehring entdeckte 1998, dass dieses Gen hochkonserviert bei allen augentragenden Tieren vorkommt.10 So gelang es mittels Maus Pax6 Augen auf den Antennen der Fruchtfliege zu induzieren und zwar Facettenaugen (Abb. 4). Natürlich sind die Regulationsgene bzw. ihre Wirkungsweise, die nach Pax6 wirken, bei den unterschiedlichen Augentypen unterschiedlich. Hier zeigt sich erneut, dass Evolution wirkt, indem sie das nimmt, was schon vorhanden ist und auf immer neue Art und Weise miteinander kombiniert.

Zum Abschluss sei noch ein Beispiel für den dritten Teilbereich von Evo-Devo angeführt. Es ist dies der dreistachlige Stichling (Gasterosteus aculeatus). Von dieser Art gibt es zwei Unterarten, eine lebt im Meer und die andere in Seen. Der marin vorkommende Stichling hat Stacheln an seinem Hüftgürtel, die der im See vorkommende Fisch nicht mehr hat (Abb. 5).11 Evolutionsbiologien argumentieren, dass der im Süßwasser lebende Fisch diese Form der Räuberabwehr wohl nicht mehr braucht. Der anatomische Unterschied zwischen diesen beiden Unterarten ist genetischer Natur. Das Gen, das für diesen Unterschied verantwortlich ist, ist Pitx1: ein Gen, das in beiden Fischen vorhanden ist, aber im benthischen Fisch nicht im Hüftgürtel expremiert wird. Es dürfte hier vor ungefähr 10.000 Jahren zu einer Mutation gekommen sein, die das Gen in der Hüfte des Stichlings stillgelegt hat. Sollte es auch im Meer zu solch einer Mutation gekommen sein, hätten die Räuber den weniger stachligen Fisch ausgerottet, indem sie ihn gefressen haben. Im See gab es solche Räuber nicht, und diese weniger wehrhafte Form konnte sich durchsetzen11 und so eine Unterart des dreistachligen Stichlings bilden.

Schlussbetrachtung
Es war ein langer Weg von den Anfängen der beiden Wissenschaften bis zum heutigen Tag. Der große Erkenntnisgewinn konnte durch die hier vorgestellten Beispiele nur angedeutet werden. Was aber zu betonen bleibt, ist die Tatsache, dass diese Wissenszunahme ohne das Zusammenarbeiten der Wissenschaftler aus beiden Gebieten nicht möglich gewesen wäre. Das Interessante an Evo-Devo ist ja, dass sich die Männer und Frauen aus dem jeweiligen Gebiet mit der Sprache und der Methodik des anderen Feldes auseinandersetzen müssen, und genau an dieser Grenzfläche zwischen zwei Gebieten ist Innovation und Fortschritt möglich und zu erwarten – etwas, das nicht nur für Evo-Devo gilt.
Referenzen
- Anthony D., The Kurgan culture. Indo-european origins and the domestication of the horse, a reconsideration, Curr Anthropol (1986); 27: 291-313
- Jacob F., Evolution and tinkering, Science (1977); 196: 1161-1166
- Wollman E. L., Jacob F., Hayes W., Conjugation and genetic recombination in Escherichia coli K-12, Cold Spring Harb Symb Quant Biol (1956); 21: 141-162
- Schimper A. F. W., Über die Entwicklung der Chlorophyllkörner und Farbkörper, Bot Z (1983); 41: 102-113
Turner S. et al., Investigating deep phylogenetic relationships among Cyanobacteria and plastids by small subunit rRNA sequence analysis, J Eukaryotic Microbiol (1999); 46: 327-338 - Garstang W., The theory of recapitulation: a critical restatement of the biogenetic law, J Linn Soc Zool (1922); 35: 81-101
- Goldschmied R. B., The Material Basis of Evolution, Yale University Press, New Haven (1940)
- Merino R. et al., The BMP antagonist Gremlin regulates outgrowth, chondrogenesis and programmed cell death in the developing limb, Devel (1999); 126: 5515-5522
- Gould. S. J., Ontogeny and Phylogeny, Harvard Univeristy Press, Cambridge (1977)
- Jacob F., Evolution and tinkering, Science (1977); 196: 1161-1166
- Gehring W. J., The master control gene for morphogenesis and evolution of the eye, Genes Cells (1996); 1(1): 11-15
- Cole N. et al., Expression of limb initiation genes and clues to the morphological diversification of threespine stickleback, Curr Biol (2003); 13: 951-952
Dr. rer. nat. Carmen Czepe
Institut für Tierzucht und Genetik
Veterinärmedizinische Universität Wien
Veterinärplatz 1, A-1210 Wien
c.czepe(at)i122server.vu-wien.ac.at



